


Eukaryote
Acerca de este escuelas selección Wikipedia
Organizar una selección Wikipedia para las escuelas en el mundo en desarrollo sin acceso a Internet era una iniciativa de SOS Children. El apadrinamiento de niños ayuda a los niños uno por uno http://www.sponsor-a-child.org.uk/ .
| Las células eucariotas Gama Temporal: hace 1,6-2100000000 años (posiblemente tan pronto como hace 2700 millones años) - Present Had'n Arcaico Proterozoico
Pha. | |
|---|---|
 | |
| Eucariotas y algunos ejemplos de su diversidad | |
| Clasificación científica | |
| Dominio: | Eucariontes Whittaker y Margulis, 1978 |
| Reinos | |
Animalia - Animales
Las amebas Plantae - Plantas Chromalveolata Rhizaria Excavata | |
| Filogenia Alternativa | |
Un eucariota (pron .: / JU k ær yo . oʊ t / EW- Karr -ee OHT-o / JU k ær yo ə t /) Es un organismo cuya células contener estructuras complejas encerrados dentro de membranas. Eucariotas más formalmente pueden ser referidos como la taxón Eukarya o eucariota. La estructura unida a la membrana que establece la definición de las células eucariotas, aparte de células procariotas es el núcleo, o envoltura nuclear, dentro del cual se lleva el material genético. La presencia de un núcleo da eucariotas su nombre, que proviene del griego ευ (eu, "bueno") y κάρυον (karyon "tuerca" o "kernel"). La mayoría de las células eucariotas también contienen otros unido a la membrana orgánulos tales como mitocondrias , cloroplastos y la Aparato de Golgi. Todos los grandes organismos complejos son eucariotas, incluyendo animales , plantas y hongos . El grupo también incluye a muchos organismos unicelulares.
La división celular en eucariotas es diferente de la de los organismos sin un núcleo ( Prokaryote). Se trata de la separación de las duplicado cromosomas, a través de movimientos dirigidos por microtúbulos. Hay dos tipos de procesos de división. En mitosis, una célula se divide para producir dos células genéticamente idénticas. En meiosis, que se requiere en la reproducción sexual, uno célula diploide (que tiene dos instancias de cada cromosoma, uno de cada padre) sufre recombinación de cada par de cromosomas parentales, y luego dos etapas de la división celular, resultando en cuatro células haploides ( gametos). Cada gameto tiene sólo un complemento de cromosomas, cada una mezcla única de la correspondiente par de cromosomas de los progenitores.
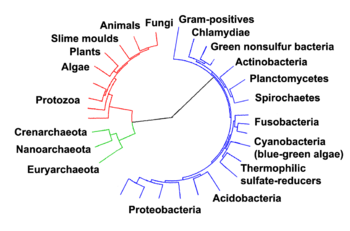
Eukaryota parece ser monofilético, y así lo hace uno de los tres dominios de la vida. Los dos dominios otros, bacterias y Archaea, son procariotas y no tienen ninguna de las características anteriores. Los eucariotas representan una pequeña minoría de todos los seres vivos; incluso en un cuerpo humano hay 10 veces más microbios que células humanas. Sin embargo, debido a su tamaño mucho más grande su biomasa colectiva en todo el mundo se estima en aproximadamente igual a la de los procariotas.
Características celulares
Las células eucariotas son típicamente mucho mayores que los de procariotas. Tienen una gran variedad de membranas y estructuras internas, llamados orgánulos, y una citoesqueleto compuesta de microtúbulos, microfilamentos, y los filamentos intermedios, que desempeñan un papel importante en la definición de la organización de la célula y forma. Eukaryotic DNA está dividido en varios paquetes lineales llamados cromosomas, que están separados por un husillo microtubular durante la división nuclear.
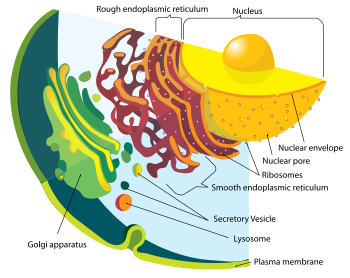
Membrana interna
Células eucariotas incluyen una variedad de estructuras unidas a la membrana, denominados colectivamente como la endomembrane sistema. Compartimientos simples, llamadas o vesículas vacuolas, pueden formar por gemación otras membranas. Muchas células ingieren los alimentos y otros materiales a través de un proceso de endocitosis, donde la membrana externa invagina y luego pellizca para formar una vesícula. Es probable que la mayoría de otros orgánulos unidos a la membrana se derivan en última instancia de tales vesículas.
El núcleo está rodeado por una membrana doble (comúnmente referido como una envoltura nuclear), con poros que permiten que el material se mueva dentro y fuera. Varias extensiones tubo- y forma de hoja de la forma de la membrana nuclear lo que se llama la retículo endoplásmico o ER, que está implicado en el transporte de proteínas y la maduración. Incluye el RE rugoso donde ribosomas se unen para sintetizar proteínas, que entran en el espacio interior o lumen. Posteriormente, por lo general entran en vesículas, que brotan fuera del RE liso. En la mayoría de los eucariotas, estas vesículas de proteínas portadoras son liberados y más modificadas en pilas de vesículas aplanadas, llamados Organismos o dictiosoma Golgi.
Las vesículas pueden ser especializadas para diversos fines. Por ejemplo, lisosomas contienen enzimas que descomponen el contenido de vacuolas de alimentos, y peroxisomas se utilizan para descomponer peróxido, que es tóxico de otra manera. Muchos protozoos tienen vacuolas contráctiles, que recogen y expulsan el exceso de agua, y extrusomes, que expulsan material utilizado para desviar los depredadores o la captura de presas. En las plantas superiores, la mayor parte del volumen de una célula está ocupada por una vacuola central, que mantiene principalmente su presión osmótica.

1) Membrana interna
2) Membrana externa
3) Crista
4) Matriz
Las mitocondrias y plastos
Las mitocondrias son orgánulos que se encuentran en casi todos los eucariotas. Están rodeados por dos membranas (cada una fosfolípido bi-capa), el interior de los cuales se dobla en invaginaciones denominadas crestas, donde respiración aeróbica tiene lugar. Las mitocondrias contienen su propio ADN. Ahora se llevan a cabo generalmente para haber desarrollado a partir de procariotas endosymbiotic, probablemente proteobacterias. Se han encontrado los unos protozoos que carecen de mitocondrias que contienen orgánulos mitocondria derivados, tales como hidrogenosomas y mitosomes; y así probablemente perdido la mitocondria secundariamente.
Las plantas y los diversos grupos de algas también tienen plástidos. Una vez más, estos tienen su propio ADN y desarrollados a partir de endosymbiotes, en este caso cianobacterias. Por lo general, toman la forma de cloroplastos, que como cianobacterias contienen clorofila y producen compuestos orgánicos (como la glucosa ) a través de la fotosíntesis . Otras participan en el almacenamiento de alimentos. Aunque plastidios probablemente tenían un origen único, no todos los grupos que contienen plastid están estrechamente relacionados. En cambio, algunos eucariotas ellas han obtenido de otros a través de la endosimbiosis secundaria o ingestión.
También se han propuesto orígenes endosymbiotic para el núcleo, para lo cual véase más adelante, y para eucariota flagelos, supone que han desarrollado a partir de espiroquetas. Esto no es generalmente aceptado, tanto de una falta de pruebas citológicas y dificultad para conciliar esto con la reproducción celular.
Estructuras del citoesqueleto
Muchos eucariotas tienen proyecciones citoplasmáticas móviles largos y delgados, llamada flagelos, o similares estructuras llamadas cilios. Flagelos y cilios se denominan a veces undulipodios, y participan de diversas formas en movimiento, la alimentación, y la sensación. Ellos se componen principalmente de tubulina. Estos son totalmente distintos de los flagelos procariota. Ellos son apoyados por un haz de microtúbulos que surgen de un basal del cuerpo, también llamado kinetosome o centríolo, característicamente dispuesto como nueve dobletes que rodean dos singletes. Los flagelos también puede tener pelos, o Mastigonema y escalas que conectan las membranas y varillas internas. Su interior es continua con la célula de citoplasma.
Estructuras compuestas por Microfilamental actina y proteínas de unión a actina, por ejemplo, α- actinina, fimbrina, filamin están presentes en las capas corticales y haces de submembraneous, también. Motor proteínas de los microtúbulos, por ejemplo, dineína o kinesin y actina, por ejemplo, myosins proporcionan carácter dinámico de la red.
Los centriolos están a menudo presentes incluso en las células y los grupos que no tienen flagelos. Por lo general, se presentan en grupos de uno o dos, llamado kinetids, que dan lugar a diversas raíces microtubulares. Estos forman un componente principal de la estructura del citoesqueleto, y a menudo se montan en el transcurso de varias divisiones celulares, con un solo flagelo retenido de la matriz y la otra derivada de ella. Los centriolos también puede estar asociada en la formación de un huso durante la división nuclear.
Importancia de las estructuras del citoesqueleto está subrayado en la determinación de la forma de las células, así como su ser componentes esenciales de las respuestas migratorias como quimiotaxis y chemokinesis. Algunos protistas tienen varios otros orgánulos microtúbulos apoyado. Estos incluyen la radiolarios y heliozoos, que producen axopodia usado en la flotación o para capturar a sus presas, y la haptophytes, que tienen un orgánulo flagelo parecido peculiar llamado haptonema.
Pared celular
Las células de plantas, hongos, y la mayoría de cromalveolados tienen una pared celular, una capa bastante rígida fuera de la membrana de la célula, proporcionando la célula con soporte estructural, la protección, y un mecanismo de filtrado. La pared celular también evita la sobre-expansión cuando el agua entra en la célula.
En las plantas, la principal polisacáridos que componen la pared celular primaria de las plantas terrestres se celulosa, hemicelulosa, y pectina. La celulosa microfibrillas están vinculados a través de correas de sujeción de hemicelulosa para formar la red de celulosa-hemicelulosa, que está incrustado en la matriz de pectina. La hemicelulosa más común en la pared celular primaria es xiloglucano.
Las diferencias entre las células eucariotas
Hay muchos tipos diferentes de células eucariotas, aunque los animales y las plantas son los eucariotas más familiares, y por lo tanto constituyen un excelente punto de partida para comprender la estructura eucariota. Los hongos y muchos protistas tienen algunas diferencias sustanciales, sin embargo.
Célula animal
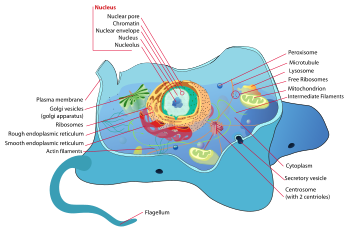
Una célula animal es una forma de célula eucariota que hace a muchos Los tejidos en los animales . La célula animal es distinto de otros eucariotas, lo más notablemente células de la planta , ya que carecen de paredes celulares y cloroplastos. También tienen menor vacuolas. Debido a la falta de un rígido la pared celular, las células animales puede adoptar una variedad de formas. La célula fagocítica incluso puede engullir otras estructuras.
Hay muchos diferentes tipos de células. Por ejemplo, hay aproximadamente 210 distintos tipos de células en el cuerpo humano adulto.
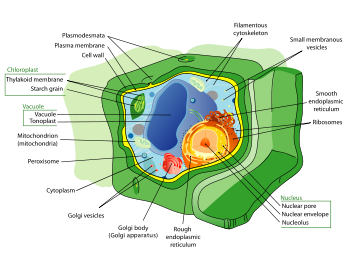
Celula vegetal
Plant células son muy diferentes de las células de los otros organismos eucariotas. Sus características son:
- Un gran centro vacuola (encerrada por una membrana, la tonoplasto), que mantiene la celda de turgencia y controla el movimiento de moléculas entre el citosol y savia
- Una primaria la pared celular que contiene celulosa, hemicelulosa y pectina, depositado por el protoplasto en el exterior de la membrana de la célula; esto contrasta con las paredes celulares de los hongos , que contienen quitina, y la envolturas celulares de procariotas, en la que peptidoglicanos son los principales moléculas estructurales
- La plasmodesmos, la vinculación de los poros en la pared celular que permiten que cada célula de la planta para comunicarse con otras células adyacentes; esto es diferente del sistema funcionalmente análogos de las uniones entre las células animales.
- Los plástidos, especialmente cloroplastos que contienen clorofila, el pigmento que da plantas su color verde y les permite realizar la fotosíntesis
- Las plantas superiores, incluyendo coníferas y plantas con flores (angiospermas) carecen de la flagelos y centriolos que están presentes en las células animales
Célula fúngica

1- hifas pared 2- 3- Septum Mitocondria 4- vacuola 5- Cristal ergosterol 6- 7- Ribosome Núcleo 8- El retículo endoplasmático cuerpo 9- 10- lípidos de membrana de plasma de 11 Aparato de Golgi Spitzenkörper 12-
Las células fúngicas son más similares a las células animales, con las siguientes excepciones:
- Una pared de celda que contiene quitina
- Menos definición entre las células; la hifas de los hongos superiores han tabiques porosos llamada septos, que permiten el paso del citoplasma, orgánulos, y, a veces, los núcleos. Hongos primitivos tienen pocos o ningún septos, por lo que cada organismo es esencialmente una supercélula multinucleadas gigantes; estos hongos se describen como coenocytic.
- Sólo los hongos más primitivo, quitridios, tienen flagelos.
Otras células eucariotas
Los eucariotas son un grupo muy diverso, y sus estructuras celulares son igualmente diversos. Muchos tienen paredes celulares; muchos no lo hacen. Muchos tienen cloroplastos, derivados de endosimbiosis primaria, secundaria, o incluso terciaria; y muchos no lo hacen. Algunos grupos tienen estructuras únicas, tales como la cyanelles de la glaucophytes, la haptonema de la haptophytes, o la ejectisomes de la cryptomonads. Otras estructuras, tales como seudópodos, se encuentran en diferentes grupos de eucariotas en diferentes formas, tales como la lobosas amoebozoans o la Reticulosa foraminíferos.
Reproducción
División nuclear a menudo se coordina con división celular. Esto generalmente se lleva a cabo por mitosis, un proceso que permite que cada núcleo hijo a recibir una copia de cada cromosoma. En la mayoría de los eucariotas, también hay un proceso de reproducción sexual, que suele implicar una alternancia entre generaciones haploides, en el que sólo una copia de cada cromosoma está presente, y generaciones diploides, en los que dos están presentes, que se producen a través de la fusión nuclear (singamia) y meiosis. Existe una variación considerable en este patrón, sin embargo.
Los eucariotas tienen una superficie más pequeña en relación al volumen de los procariotas, y por lo tanto tienen tasas metabólicas más bajas y tiempos de generación más largos. En algunos organismos multicelulares, células especializadas para el metabolismo se han ampliado áreas de superficie, tales como vili intestinales.
Clasificación

Incluso de vuelta a la Antigüedad los dos clados de animales y plantas fueron reconocidos. Se les dio la rango taxonómico de Unido (biología) por Linneo . A pesar de que incluía los hongos con plantas con algunas reservas, más tarde se dio cuenta de que son muy diferentes y requieren un reino independiente, cuya composición no estaba del todo claro hasta la década de 1980. Los diversos eucariotas unicelulares se colocaron originalmente con plantas o animales cuando se hicieron conocidos. El biólogo alemán Georg A. Goldfuss acuñó la palabra protozoos en 1830 para referirse a organismos como ciliados y corales, y este grupo se amplió hasta que abarcaba todos los eucariotas unicelulares, y dado su propio reino, el Protista por Ernst Haeckel en 1866. Las eucariotas así llegó a estar compuesto por cuatro reinos:
Los protistas se entiende que son "formas primitivas", y por lo tanto una grado evolutivo, unidos por su naturaleza primitiva unicelular. El desenredo de las profundas divisiones en la árbol de la vida en realidad sólo se puso en marcha con La secuenciación del ADN, dando lugar a un sistema de dominios en lugar de reinos como el rango de nivel superior que se presentaron por Carl Woese, uniendo a todos los reinos eucariotas bajo el dominio eucariota. Al mismo tiempo, el trabajo en el árbol protista se intensificó, y aún prosigue de forma regular en la actualidad. Varias clasificaciones alternativas se han remitido, aunque no existe un consenso en el campo.
La clasificación producida en 2005 por la Sociedad Internacional de Protistologists, que refleja el consenso de las veces, divide los eucariotas en seis "supergrupos" supuestamente monofiléticos. Aunque la clasificación publicada deliberadamente no hizo uso de rangos taxonómicos formales, otras fuentes han tratado a cada uno de los seis como reino independiente.
| Excavata | Vario protozoos flagelados |
| Amoebozoa | La mayoría lobosas ameboides y moldes de limo |
| Opisthokonta | Animales , hongos , coanoflagelados, etc. |
| Rhizaria | Foraminíferos, Radiolaria, y varios otros protozoos ameboides |
| Chromalveolata | Stramenopiles (o Heterokonta), Haptophyta, Cryptophyta (o cryptomonads), y Alveolata |
| Archaeplastida (o Primoplantae) | Las plantas terrestres, algas verdes, algas rojas, y glaucophytes |
Sin embargo, en el mismo año (2005), se expresaron dudas sobre si algunos de estos supergrupos eran monofilético, en particular el Chromalveolata, y una revisión en 2006 señalaron la falta de pruebas para varios de los supuestos seis supergrupos.
Filogenia
árboles rRNA construidos durante los años 1980 y 1990 dejaron la mayoría de los eucariotas en un grupo de "corona" sin resolver (no técnicamente un verdadero corona), que por lo general se dividió por la forma de las crestas mitocondriales; ver eucariotas corona. Los pocos grupos que carecen de mitocondrias ramificado separado, y así la ausencia se creía que era primitiva; pero esto ahora se considera un artefacto de larga rama atracción, y que se sabe que han perdido secundariamente.
A partir de 2011, existe un acuerdo generalizado de que el Rhizaria pertenecen con Estramenópilos y la Alveolata, en un clado denominado el SAR supergrupo, de modo que Rhizara no es uno de los principales grupos de eucariotas; también que el Amoeboza y Opisthokonta son cada monofilético y forman un clado, a menudo llamado el unikonts. Más allá de esto, no parece haber un consenso.
Se ha estimado que puede haber 75 linajes distintos de los eucariotas. La mayoría de estos linajes son protistas.
Los tamaños del genoma eucariota conocidos varían de 8,2 megabases (Mb) en Babesia bovis a 112,000-220,050 Mb en el dinoflagelado Micans Prorocentrum que sugieren que el genoma de la eucariota ancestral ha sufrido variación considerable durante su evolución. El último ancestro común de todos los eucariotas se cree que ha sido un protista phagotrophic con un núcleo, al menos un centríolo y cilio, mitocondria aeróbica facultativamente, el sexo ( meiosis y singamia), un estado latente quiste con una pared celular de quitina y / o celulosa y peroxisomas. Más tarde endosimbiosis llevado a la propagación de plástidos en algunos linajes.
Chromalveolata + Rhizaria
Algunos análisis desmontar la Chromalveolata + Rhizaria, mostrando una estrecha relación con la Archaeplastida. Por ejemplo, en 2007, Burki et al. Produce un árbol de la forma que se muestra a continuación.
| |||||||||||||||||||||||||||||||||||||||||||
Bikonts y unikonts
En otro análisis, el Hacrobia se muestran anidado dentro del Archaeplastida, que en conjunto forman un clado con la mayor parte del Excavata, antes de unirse al clado SAR de Stramenopiles, Alveolata y Rhizaria. Junto a todos estos grupos constituyen la bikonts, la Amoebozoa y Opisthokonta la formación de la unikonts.
| ||||||||||||||||||||||||||||||||||||||||
La división de los eucariotas en dos clados primarios, unikonts y bikonts, derivada de un organismo ancestral uniflagellar y un organismo biflagellar ancestral, respectivamente, se ha sugerido antes.
Un estudio de 2012 produjo una división algo similar, aunque señalar que los términos "unikonts" y "bikonts" no se utilizaron en el sentido original.
| eucariotas |
| ||||||||||||||||||||||||||||||
Ampliado Chromalveolata
Otros análisis lugar el supergrupo SAR dentro de un Chromalveolata expandido, aunque difieren en la colocación de los cinco grupos resultantes. Rogozin et al., En 2009 produjo el árbol se muestra a continuación, donde la división principal es entre el Archaeplastida y todos los otros eucariotas.
| |||||||||||||||||||||||||||||||||||||
Más comúnmente la expandido Chromalveolata se muestra como más estrechamente relacionado con el Archaeplastida, produciendo un árbol de la forma mostrada a continuación.
| |||||||||||||||||||||||||||||||||||||
Puntos de vista alternativos
Un artículo publicado en 2009, que reexaminada de los datos utilizados en algunos de los análisis presentados anteriormente, así como la realización de otras nuevas, sugiere fuertemente que la Archaeplastida son polifilético. La filogenia finalmente propuesto en el documento se muestra a continuación.
| eucariotas |
| |||||||||||||||||||||||||||||||||
También hay pequeños grupos de eucariotas - incluyendo el género Collodictyon, la telonemids y biliphytes - cuya posición es incierta o parece caer fuera de los grupos principales. En general, parece que, si bien se han logrado avances, aún existen dudas muy importantes en la historia de la evolución y la clasificación de los eucariotas. Como dijo Roger & Simpson en 2009 ", con el ritmo actual de cambio en nuestra comprensión del árbol eucariota de la vida, debemos proceder con cautela."
Origen de los eucariotas

El origen de la célula eucariota se considera un hito en la evolución de la vida, ya que incluyen todas las células complejas y casi todos los organismos multicelulares. El momento de esta serie de eventos es difícil de determinar; Knoll (2006) sugiere que desarrollaron hace aproximadamente 1,6 hasta 2100 millones de años. Algunos acritarcos se conocen desde al menos 1650 millones de años atrás, y la posible alga Grypania ha encontrado ya en hace 2.1 mil millones años.
Estructuras vivas organizadas han sido encontrados en lutitas negras de la Formación Paleoproterozoica Francevillian B en Gabón, fechadas en 2,1 mil millones de años. La vida eucariota podría haber evolucionado en ese momento. Los fósiles que están claramente relacionados con grupos modernos comienzan a aparecer alrededor de 1,2 millones de años, en la forma de una alga roja, aunque trabajos recientes sugieren la existencia de fosilizada algas filamentosas en el Vindhya cuenca remonta quizás a hace 1,6 hasta 1,7 mil millones de años.
Biomarcadores sugieren que al menos derivan eucariotas surgieron incluso antes. La presencia de esteranos en Australia pizarras indica que los eucariotas estaban presentes en estas rocas fechadas en 2,7 mil millones de años.
Relación con Archaea
Los eucariontes están más estrechamente relacionados con Las arqueas de las bacterias , al menos en términos de ADN nuclear y maquinaria genética, y una idea controversial es colocarlos con Archaea en el clado Neomura. Sin embargo, en otros aspectos, tales como la composición de la membrana, eucariotas son similares a las bacterias. Se han propuesto tres explicaciones principales para esto:
- Eucariotas el resultado de la fusión completa de dos o más células, en el que el citoplasma forma a partir de un eubacterium, y el núcleo de un archaeon, a partir de un virus, o de una pre-célula.
- Los eucariotas desarrollaron a partir de las arqueas, y adquirieron sus características eubacterianas del proto-mitocondria.
- Eucariotas y Archaea desarrollaron por separado de un eubacterium modificado.
También existe la teoría Kronocyte para el origen de la célula eucariota. Esta postula que una célula eucariota primitiva surgió del mundo pre-ADN pero conservó la química basada en ARN anterior de la que surgió la vida moderna. Esta célula primitiva se llama el Kronocyte. De acuerdo con esta hipótesis un Kronocyte basada en ARN coexistió con la basada en el ADN Archaea (y probablemente eubacterias) y se convirtió en la célula eucariota moderna después de una serie de importantes endosymbioses-la primera fue la incorporación de un Archaea que introdujo el metabolismo del ADN y el núcleo, a continuación, la incorporación de un alphaproteobacter que se convirtió en la mitocondria (y bacterias fotosintéticas encuentran en las plantas de hoy en día como cloroplastos). La hipótesis Kronocyte explica el gran número de genes que son hoy en día sólo se encuentran en eucariotas, pero no en Archaea o bacterias.
Sistema Endomembrane y las mitocondrias
Los orígenes del sistema endomembrane y mitocondrias son también claras. La hipótesis phagotrophic propone que las membranas de tipo eucariota que carecen de una pared celular se originaron primero, con el desarrollo de la endocitosis, mientras que las mitocondrias fueron adquiridas por ingestión como endosimbiontes. La hipótesis syntrophic propone que el proto-eucariota se basó en el proto-mitocondria por la comida, por lo que en última instancia, creció a rodearlo. Aquí las membranas se originaron después de la inmersión de la mitocondria, en parte gracias a los genes mitocondriales (el hipótesis de hidrógeno es una versión particular).
En un estudio con los genomas de construir supertrees, Pisani et al. (2007) sugieren que, junto con la evidencia de que nunca hubo un eucariota-mitocondria menos, eucariotas evolucionaron a partir de un syntrophy entre un arqueas estrechamente relacionado con Thermoplasmatales y un α-proteobacteria, probablemente una simbiosis impulsado por azufre o hidrógeno. La mitocondria y su genoma es un remanente de la endosymbiont-α proteobacterial.
Las hipótesis sobre el origen de los eucariotas
Diferente hipótesis han sido propuestas en cuanto a cómo las células eucariotas llegaron a existir. Estas hipótesis se pueden clasificar en dos clases distintas - autógenos modelos y modelos quiméricos.
Modelos autógena
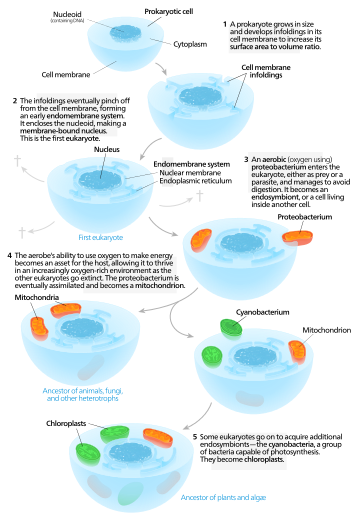 |
| Un modelo autógena para el origen de los eucariotas. |
Modelos autógena proponen que una célula originalmente proto-eucariota que contiene una existía núcleo primero, y más tarde adquirió mitocondrias . De acuerdo con este modelo, una gran procariotas desarrollado invaginaciones en su membrana plasmática con el fin de obtener suficiente área de la superficie para el servicio de su volumen citoplasmático. Como las invaginaciones diferenciados en función, algunos se convirtieron en independientes compartimentos que da lugar a la endomembrane sistema, incluyendo la retículo endoplásmico, aparato de golgi, la membrana nuclear, y sola membrana estructuras tales como lisosomas. Las mitocondrias se propuso venir del endosimbiosis de una aeróbico proteobacteria, y se asume que todos los linajes eucariotas que no adquirieron las mitocondrias se extinguieron. Los cloroplastos surgió de otro evento que involucra endosimbiótica cianobacterias. Dado que todos los eucariotas tienen mitocondrias, pero no todas tienen cloroplastos, mitocondrias se cree haber llegado primero. Este es el teoría de la endosimbiosis serial.
Algunos modelos proponen que los orígenes de doble capa orgánulos tales como mitocondrias y cloroplastos en la célula eucariota es proto-debido a la compartimentación de ADN vesículas que se forma a partir de las invaginaciones secundarias o repliegues más detalladas de la membrana celular.
Modelos quiméricos
Modelos quiméricos afirman que dos células procariotas existían inicialmente - una archaeon y una bacteria . Estas células se sometieron a un proceso de fusión, ya sea por una fusión física o por endosimbiosis, lo que conduce a la formación de una célula eucariota. Dentro de estos modelos quiméricos, algunos estudios afirman, además, que las mitocondrias se originó a partir de un antepasado bacteriano, mientras que otros hacen hincapié en el papel de los procesos endosymbiotic detrás del origen de las mitocondrias.
Basado en el proceso de simbiosis mutualista, las hipótesis pueden ser categorizadas como - la serie teoría endosimbiótica (SET), la hipótesis de hidrógeno (principalmente un proceso de simbiosis donde la transferencia de hidrógeno se lleva a cabo entre las diferentes especies), y la hipótesis syntrophy.
Según la teoría de la endosimbiosis serial, la unión entre un motile bacteria anaeróbica (como Spirochaeta) y un crenarchaeon termoacidofílicos (como Thermoplasma que es sulfidogénicos en la naturaleza) dieron origen a las presentes eucariotas día. Esta unión estableció un organismo móviles capaces de vivir en las aguas ácidas y sulfurosas ya existentes. El oxígeno se sabe que causa toxicidad para los organismos que carecen de la requeridos maquinaria metabólica. Así, el archaeon proporcionó la bacteria con un entorno altamente beneficioso reducida (azufre y sulfato se reducen a sulfuro). En condiciones microaerofílicas, el oxígeno se redujo al agua creando así una plataforma de beneficio mutuo. La bacteria en el otro lado, aportó el necesario productos de fermentación y electrones aceptantes junto con su característica motilidad al archaeon obteniendo con ello una piscina la motilidad para el organismo. A partir de un consorcio de ADN bacteriano y arqueas originado la nuclear genoma de las células eucariotas. Las espiroquetas dieron lugar a las características de movilidad de las células eucariotas. Unificaciones endosimbióticas de los antepasados de la alfa-proteobacterias y cianobacterias, llevaron al origen de las mitocondrias y plastidios respectivamente. Por ejemplo, Thiodendron se ha sabido que se originó a través de un ectosymbiotic proceso basado en un similares syntrophy de azufre existente entre los dos tipos de bacterias - Desulphobacter y Spirochaeta. Sin embargo, tal asociación basado en simbiosis móviles nunca se han observado en la práctica. Además, no hay evidencia de archaea y espiroquetas adaptándose a entornos basados en ácidos intensos.
En la hipótesis de hidrógeno, la relación simbiótica de un anaeróbico y autotrófica archaeon metanogénica (host) con un-proteobacteria alfa (el simbionte) dio lugar a los eucariotas. El huésped utilizado de hidrógeno (H 2) y dióxido de carbono (CO 2) para producir metano mientras que el simbionte, capaz de la respiración aeróbica, expulsado H 2 y CO 2 como subproductos del proceso de fermentación anaerobia. Entorno metanogénica del anfitrión trabajó como sumidero de H 2, que resultó en la fermentación bacteriana aumentada. La transferencia de genes Endosymbiotic (EGT) actuó como un catalizador para el anfitrión para adquirir los simbiontes ' carbohidratos metabolismo y turno heterótrofos en la naturaleza. Posteriormente, la formación de metano capacidad del huésped se perdió. Por lo tanto, el origen de los orgánulos heterótrofos (simbionte) son idénticos a los orígenes de los eucariotas linaje. En esta hipótesis, la presencia de H 2 representa la fuerza selectiva que forjó eucariotas de procariotas.
La hipótesis syntrophy fue desarrollado en contraste con la hipótesis de hidrógeno y propone la existencia de dos eventos simbióticas. Según esta teoría, eukaryogenesis (es decir, el origen de las células eucariotas) se produjo basa en simbiosis metabólica (syntrophy) entre una archaea metanogénicas y un delta-proteobacteria. Esta simbiosis syntrophic fue facilitada inicialmente por H 2 transferencia entre diferentes especies en ambientes anaeróbicos. En las primeras etapas, un alfa-proteobacteria convirtió en miembro de esta integración, y más tarde se convirtió en la mitocondria. La transferencia de genes de una delta-proteobacteria a un archaeon llevó a la archaea metanogénicas convertirse en un núcleo. El archaeon constituía el aparato genético mientras que el delta-proteobacteria contribuyó a la características citoplasmática. Esta teoría incorpora dos fuerzas selectivas que se necesitaban para ser considerado en la época del núcleo evolución - (a) la presencia de compartimentación metabólica con el fin de evitar los efectos nocivos de la co-existencia de anabólicos y vías celulares catabólicas, y (b) la prevención de anormal la biosíntesis de las proteínas que se producen debido a una amplia propagación de intrones en los genes de arqueas después de la adquisición de la mitocondria y la pérdida de metanogénesis.
Así, el origen de los eucariotas mediante procesos de endosymbiotic ha sido ampliamente reconocido y aceptado hasta ahora. Las mitocondrias y plastidios se ha sabido que se originan a partir de un antepasado bacteriano durante la adaptación paralela a la anaerobiosis. Sin embargo, todavía existe una necesidad mayor en la evaluación de la cuestión de cuánto complejidad eucariota se está originada a través de una implementación de estos teorías symbiogenetic.