

Evolución
Acerca de este escuelas selección Wikipedia
Esta selección Escuelas fue originalmente elegido por SOS para las escuelas en el mundo en desarrollo que no tienen acceso a Internet. Está disponible como una descarga intranet. Con Infantil SOS se puede elegir a apadrinar a los niños en más de cien países
La evolución es el cambio en el heredado características de biológica poblaciones más sucesiva generaciones. Los procesos evolutivos dan lugar a la diversidad en todos los nivel de organización biológica, incluyendo las especies , los organismos individuales y moléculas tales como ADN y proteínas .
La vida en la Tierra evolucionó a partir de un ancestro común universal hace aproximadamente 3,8 millones de años. Repetido especiación y la divergencia de la vida puede ser inferirse de conjuntos compartidos de caracteres bioquímicos y morfológicos, o por compartieron Secuencias de ADN. Estos rasgos y secuencias homólogas son más similares entre las especies que comparten un ancestro común más reciente, y se pueden utilizar para reconstruir historias evolutivas, usando ambas especies existentes y el registro fósil . Los patrones existentes de la biodiversidad han sido en forma de tanto por la especialización y por la extinción .
Charles Darwin y Alfred Wallace fueron los primeros en formular un argumento científico para la teoría de la evolución mediante la selección natural . La evolución por selección natural es un proceso que se infiere de tres hechos sobre las poblaciones: 1) más descendencia se producen los que pueden sobrevivir, 2) rasgos varían entre los individuos, lo que lleva a diferentes tasas de supervivencia y reproducción, y 3) las diferencias de rasgos son hereditarias. Por lo tanto, cuando los miembros de una población mueren son reemplazados por la progenie de padres que eran mejores adaptado para sobrevivir y reproducirse en el entorno en el que la selección natural tuvo lugar. Este proceso crea y preserva los rasgos que son aparentemente equipada para el roles funcionales que realizan. La selección natural es la única causa conocida de adaptación, pero no la única causa conocida de la evolución. Otras causas, no adaptativas de evolución incluye mutación y deriva genética.
En el siglo 20, la genética era integrado con la teoría de la evolución por selección natural de Darwin a través de la disciplina de la genética de poblaciones. La importancia de la selección natural como una de las causas de la evolución fue aceptada en otras ramas de la biología . Nociones Además, celebradas con anterioridad acerca de la evolución, como ortogénesis y "Progreso" se convirtió en obsoleto. Los científicos continúan estudiar diversos aspectos de la evolución mediante la formación y comprobar hipótesis, construir las teorías científicas, utilizando datos de observación y rendimiento experimentos tanto en el campo y el laboratorio. Biólogos de acuerdo en que la descendencia con modificación es uno de lo establecido más fiable hechos en la ciencia. Los descubrimientos en la biología evolutiva han tenido un impacto significativo no sólo dentro de las ramas tradicionales de la biología, sino también en otras disciplinas académicas (por ejemplo, la antropología y la psicología) y en la sociedad en general.
Historia del pensamiento evolutivo
La propuesta de que un tipo de animal podría descender de un animal de otro tipo se remonta a algunos de los primeros filósofos griegos presocráticos, como Anaximandro y Empédocles. En contraste con estos puntos de vista materialista, Aristóteles entienden todas las cosas naturales, no sólo viven las cosas, como ser imperfecto actualisations de diferentes posibilidades naturales fijos, conocidos como " formas "," ideas ", o (en las traducciones latinas)" especies ". Esta fue parte de su comprensión teleológica de naturaleza en la que todas las cosas tienen un papel destinado a jugar en un divino orden cósmico. La Romana poeta y filósofo Tito Lucrecio Caro propuso la posibilidad de cambios evolutivos de los organismos. Las variaciones de esta idea se convirtió en la comprensión estándar de la Edad Media , y se integraron en el aprendizaje cristiano, pero Aristóteles no exigen que los tipos reales de animales corresponden uno a uno con las formas metafísicas exactas, y en concreto dio ejemplos de cómo los nuevos tipos de los seres vivos podría llegar a ser.
En el siglo 17 el nuevo método de la ciencia moderna rechazó el planteamiento de Aristóteles, y buscó explicaciones de los fenómenos naturales en términos de leyes de la naturaleza, que eran los mismos para todas las cosas visibles, y no necesitan asumir ninguna categoría naturales fijos, ni ninguna orden cósmico divino. Pero este nuevo enfoque fue lento a echar raíces en las ciencias biológicas, que se convirtió en el último bastión del concepto de tipos naturales fijos. John Ray utiliza uno de los términos previamente más generales para tipos naturales fijos, "especie", que aplicará a los tipos de animales y plantas, pero a diferencia de Aristóteles que estrictamente identifica cada tipo de ser viviente como especie, y propuso que cada especie se pueden definir por las características que perpetúan a sí mismos cada generación. Estas especies fueron diseñados por Dios, sino que muestran las diferencias causadas por las condiciones locales. La clasificación biológica introducida por Carlos Linneo en 1735 también vieron especies como fijo de acuerdo a un plan divino.

Otros naturalistas de este tiempo especularon sobre el cambio evolutivo de las especies a través del tiempo de acuerdo con las leyes naturales. Maupertuis escribió en 1751 de las modificaciones naturales que ocurren durante la reproducción y acumulación durante muchas generaciones para producir nuevas especies. Buffon sugirió que las especies podrían degenerar en diferentes organismos, y Erasmus Darwin propuso que todos los animales de sangre caliente podrían haber descendido de un solo microorganismo (o "filamento"). El primer esquema evolutivo de pleno derecho fue Teoría de la "transmutación" de Lamarck de 1809, que preveía la generación espontánea produciendo continuamente formas simples de vida desarrolló una mayor complejidad en linajes paralelos con una tendencia progresiva inherente, y que a nivel local estos linajes adaptado al medio ambiente por los cambios que heredan causados por el uso o desuso en padres. (Este último proceso se llamó más tarde lamarckismo.) Estas ideas fueron condenados por los naturalistas establecidos como la especulación carente de sustento empírico. En particular Georges Cuvier insistió en que las especies no estaban relacionados y fija, sus similitudes que refleja el diseño divino para las necesidades funcionales. Mientras tanto, las ideas de Ray de diseño benevolente habían sido desarrollados por William Paley en un teología natural que propone adaptaciones complejas como evidencia de designio divino, y fue admirado por Charles Darwin.
La ruptura crítica del concepto de especies fijas en biología comenzó con la teoría de la evolución por selección natural, que fue formulada por Charles Darwin. Parcialmente influenciado por Ensayo sobre el principio de la población por Thomas Robert Malthus , Darwin observó que el crecimiento de la población llevaría a una "lucha por la existencia", donde las variaciones favorables podrían prevalecer como otros perecieron. Cada generación, muchos hijos no logran sobrevivir a la era de la reproducción debido a los recursos limitados. Esto podría explicar la diversidad de animales y plantas a partir de un ancestro común por la acción de las leyes naturales que trabajan el mismo para todos los tipos de cosas. Darwin estaba desarrollando su teoría de la " selección natural "de 1838 en adelante hasta Alfred Russel Wallace le envió una teoría similar en 1858. Ambos hombres presentaron su documentos separados a la Linnean Society de Londres. A finales de 1859, la publicación de Darwin de El Origen de las Especies , explicó la selección natural en detalle y de una manera que condujo a una aceptación cada vez más amplia de La evolución darwiniana. Thomas Henry Huxley aplicó las ideas de Darwin a los seres humanos, mediante la paleontología y anatomía comparada para proporcionar una fuerte evidencia de que los humanos y los simios comparten un ancestro común. Algunos fueron perturbados por esto, ya que implica que los humanos no tienen un lugar especial en el universo.
Mecanismos precisos de heredabilidad reproductiva y el origen de nuevos rasgos seguían siendo un misterio. Con este fin, Darwin desarrolló su teoría provisional de pangénesis. En 1865 Gregor Mendel informó que los rasgos se heredan de una manera predecible a través de la distribución independiente y segregación de los elementos (más tarde conocido como genes). Las leyes de la herencia de Mendel finalmente suplantados más de la teoría de Darwin pangénesis. August Weismann hizo la importante distinción entre las células germinales (espermatozoides y óvulos) y las células somáticas del cuerpo, lo que demuestra que la herencia pasa a través de sólo la línea germinal. Hugo de Vries conectado teoría de Darwin a pangénesis germen / distinción célula soma de Weismann y propuso que pangenes de Darwin se concentraron en el núcleo de la célula y cuando se expresa que pudieran moverse en el citoplasma para cambiar la estructura de las células. De Vries fue también uno de los investigadores que hicieron el trabajo de Mendel conocido, en la creencia de que los rasgos mendelianos correspondieron a la transferencia de las variaciones heredables a lo largo de la línea germinal. Para explicar cómo se originan nuevos variantes, De Vries desarrolló un teoría de la mutación que llevó a una ruptura temporal entre los que aceptaban la evolución darwiniana y biometristas que alió con de Vries. A la vuelta del siglo 20, pioneros en el campo de la genética de poblaciones, tales como JBS Haldane, Sewall Wright, y Ronald Fisher, sentó las bases de la evolución hacia una filosofía estadístico robusto. La falsa contradicción entre la teoría de Darwin, las mutaciones genéticas, y la herencia mendeliana fue así reconciliarse.
En la década de 1920 y 1930 un la síntesis evolutiva moderna conectada la selección natural, la teoría de la mutación, y la herencia mendeliana en una teoría unificada que aplica generalmente a cualquier rama de la biología. La síntesis moderna fue capaz de explicar los patrones observados en todas estas especies en las poblaciones, a través de transiciones fósiles en la paleontología, y los mecanismos celulares más complejas en la biología del desarrollo. La publicación de la estructura del ADN por James Watson y Francis Crick en 1953 demostró una base física para la herencia. La biología molecular ha mejorado nuestra comprensión de la relación entre genotipo y fenotipo. Los avances también se hicieron en filogenético la sistemática, la cartografía de la transición de los rasgos en un marco comparativo y comprobable a través de la publicación y el uso de árboles evolutivos. En 1973, el biólogo evolutivo Theodosius Dobzhansky escribió que "nada en biología tiene sentido excepto a la luz de la evolución", ya que ha sacado a la luz las relaciones de lo que primero parecía hechos inconexos de la historia natural en un coherente cuerpo explicativo de conocimiento que describe y predice muchos hechos observables acerca de la vida en este planeta.
Desde entonces, la síntesis moderna se ha extendido más allá para explicar los fenómenos biológicos a través de la escala completa e integradora de las jerarquía biológica, a partir de genes de especies. Esta extensión ha sido apodado " eco-evo-devo ".
Herencia
Evolución en los organismos se produce a través de cambios en heredable rasgos - características particulares de un organismo. En los seres humanos, por ejemplo, color de ojos es una característica heredada y un individuo puede heredar el "rasgo de ojos marrón" de uno de sus padres. Los rasgos hereditarios son controlados por genes y el conjunto completo de genes dentro de un organismo de genoma se llama su genotipo.
El conjunto completo de características observables que componen la estructura y el comportamiento de un organismo se denomina su fenotipo. Estos rasgos proceden de la interacción de su genotipo con el medio ambiente. Como resultado, no se heredan muchos aspectos de fenotipo de un organismo. Por ejemplo, piel bronceada proviene de la interacción entre el genotipo y la luz del sol de una persona; por lo tanto, el bronceado no se transmiten a los hijos de las personas. Sin embargo, algunas personas se broncean más fácilmente que otros, debido a las diferencias en su genotipo; un claro ejemplo son las personas con el rasgo heredado de albinismo , que no lo hacen tan en absoluto y son muy sensibles a quemaduras solares.
Rasgos hereditarios se transmiten de una generación a otra a través del ADN , una molécula que codifica la información genética. El ADN es un largo polímero compone de cuatro tipos de bases. La secuencia de bases a lo largo de una molécula de ADN particular, especificar la información genética, de una manera similar a una secuencia de letras deletrear una frase. Antes de que una célula se divide, se copia el ADN, de modo que cada una de las dos células resultantes heredarán la secuencia de ADN. Las porciones de una molécula de ADN que especifican una sola unidad funcional se denominan genes; diferentes genes tienen diferentes secuencias de bases. Dentro de las células , los largos mechones de forma condensada estructuras de ADN llamados cromosomas. La ubicación específica de una secuencia de ADN dentro de un cromosoma se conoce como una locus. Si la secuencia de ADN en un locus varía entre los individuos, las diferentes formas de esta secuencia se denominan alelos. Secuencias de ADN pueden cambiar a través mutaciones, la producción de nuevos alelos. Si se produce una mutación dentro de un gen, el nuevo alelo puede afectar el rasgo de que los controles de genes, alterando el fenotipo del organismo. Sin embargo, mientras que este simple correspondencia entre un alelo y un rasgo funciona en algunos casos, la mayoría de los rasgos son más complejos y son controlados por múltiples genes que interactúan.
Hallazgos recientes han confirmado importantes ejemplos de cambios heredables que no se pueden explicar por los cambios a la secuencia de nucleótidos en el ADN. Estos fenómenos se clasifican como sistemas de herencia epigenética. La metilación del ADN marcado cromatina, bucles metabólicas auto-sostenible, silenciamiento génico por La interferencia de ARN y el tridimensional conformación de proteínas (tales como priones ) son áreas donde los sistemas de herencia epigenética han sido descubiertos en el nivel orgánico. Los biólogos del desarrollo sugieren que las interacciones complejas en redes genéticas y la comunicación entre las células pueden dar lugar a variaciones heredables que pueden UNDERLAY algunas de las mecánicas en plasticidad del desarrollo y canalización. La heredabilidad también puede ocurrir a escala aún mayor. Por ejemplo, la herencia ecológica a través del proceso de construcción nicho se define por las actividades regulares y repetidas de organismos en su medio ambiente. Esto genera un legado de los efectos que modifican y se alimentan de nuevo en el régimen de selección de las generaciones posteriores. Descendientes heredan genes más características ambientales generados por las acciones ecológicas de los antepasados. Otros ejemplos de heredabilidad en evolución que no están bajo el control directo de genes incluyen la herencia de rasgos culturales y simbiogénesis.
Variación


De un organismo individual resultados fenotipo tanto de su genotipo y la influencia de la medio ambiente que ha vivido en. Una parte sustancial de la variación en los fenotipos en una población es causada por las diferencias entre sus genotipos. La la síntesis evolutiva moderna define la evolución como el cambio en el tiempo de esta variación genética. La frecuencia de un alelo particular, se hará más o menos frecuente en relación con otras formas de ese gen. Variación desaparece cuando un nuevo alelo alcanza el punto de fijación - cuando sea desaparece de la población o reemplaza el alelo ancestral por completo.
La selección natural sólo hará que la evolución, si hay suficiente la variación genética en una población. Antes del descubrimiento de La genética mendeliana, una hipótesis común era herencia de mezcla. Pero con la herencia mezclada, la variación genética se pierde rápidamente, por lo que la evolución por selección natural inverosímil. La Principio de Hardy-Weinberg ofrece la solución a cómo se mantiene la variación en una población con Herencia mendeliana. Las frecuencias de los alelos (variaciones en un gen) se mantendrá constante en ausencia de selección, mutación, la migración y la deriva genética.
Variación proviene de mutaciones en material genético, reorganización de los genes a través de la reproducción sexual y la migración entre poblaciones ( flujo de genes). A pesar de la constante introducción de nueva variación través de la mutación y el gen de flujo, la mayoría de las genoma de una especie es idéntico en todos los individuos de dicha especie. Sin embargo, incluso relativamente pequeñas diferencias en el genotipo pueden llevar a diferencias dramáticas en el fenotipo: por ejemplo, los chimpancés y los seres humanos difieren en sólo alrededor del 5% de sus genomas.
Mutación
Las mutaciones son cambios en la secuencia de ADN del genoma de una célula. Cuando se producen mutaciones, que pueden o bien no tienen ningún efecto, alterar el producto de un gen, o prevenir que el gen funcione. Con base en estudios en la mosca Drosophila melanogaster , se ha sugerido que si una mutación cambia una proteína producida por un gen, esto probablemente será perjudicial, con aproximadamente 70% de estas mutaciones que tienen efectos perjudiciales, y siendo el resto neutro o débilmente beneficioso.
Las mutaciones pueden implicar grandes secciones de un cromosoma convertirse duplicado (generalmente por recombinación genética), que puede introducir copias adicionales de un gen en un genoma. Copias adicionales de genes son una fuente importante de la materia prima necesaria para los nuevos genes a evolucionar. Esto es importante porque la mayoría de los nuevos genes evolucionan dentro de familias de genes de genes preexistentes que comparten antepasados comunes. Por ejemplo, el ojo humano utiliza cuatro genes para que las estructuras que detectan la luz: tres para la visión del color y uno para vision nocturna; los cuatro son descendientes de un único gen ancestral.
Nuevos genes pueden ser generados a partir de un gen ancestral cuando un duplicado copia muta y adquiere una nueva función. Este proceso es más fácil una vez que un gen ha sido duplicado, ya que aumenta la redundancia del sistema; un gen en el par puede adquirir una nueva función, mientras que la otra copia sigue llevando a cabo su función original. Otros tipos de mutaciones pueden incluso generar completamente nuevos genes de ADN no codificante previamente.
La generación de nuevos genes también puede implicar pequeñas partes de varios genes se duplican, con estos fragmentos a continuación, recombinar para formar nuevas combinaciones con nuevas funciones. Cuando los nuevos genes son ensambladas a partir barajar partes preexistentes, dominios actúan como simples módulos con funciones independientes, que se pueden mezclar juntos para producir nuevas combinaciones con funciones nuevas y complejas. Por ejemplo, policétido sintasas son enzimas grandes que hacen que los antibióticos; que contienen hasta cien dominios independientes que cada catalizar un paso en el proceso general, como un paso en una línea de montaje.
Sexo y recombinación
En los organismos asexuales, los genes se heredan juntos, o vinculados, ya que no pueden mezclarse con genes de otros organismos durante la reproducción. En contraste, la descendencia de organismos sexuales contienen mezclas aleatorias de los cromosomas de los padres que se producen a través de distribución independiente. En un proceso relacionado llamado recombinación homóloga, los organismos sexuales intercambiar ADN entre dos cromosomas juego. La recombinación y redistribución no alteran las frecuencias de alelos, pero en vez cambian alelos que están asociados entre sí, produciendo con nuevas combinaciones de alelos descendencia. Sexo por lo general aumenta la variación genética y puede aumentar la tasa de evolución.
El flujo de genes
El flujo de genes es el intercambio de genes entre poblaciones y entre las especies. Por lo tanto, puede ser una fuente de variación que es nuevo para una población o para una especie. El flujo de genes puede ser causada por el movimiento de individuos entre poblaciones separadas de los organismos, como podría ser causado por el movimiento de los ratones entre el interior y las poblaciones costeras, o el movimiento de polen entre tolerante heavy metal y heavy metal poblaciones sensibles de hierbas.
La transferencia de genes entre especies incluye la formación de organismos híbridos y transferencia horizontal de genes. La transferencia génica horizontal es la transferencia de material genético de un organismo a otro organismo que no es su descendencia; esto es más común entre las bacterias . En la medicina, esto contribuye a la propagación de resistencia a los antibióticos, como cuando una bacteria adquiere genes de resistencia se puede transferir rápidamente a ellos otras especies. La transferencia horizontal de genes desde las bacterias hasta los eucariotas tales como la levadura Saccharomyces cerevisiae y la adzuki frijol escarabajo Callosobruchus chinensis se ha producido. Un ejemplo de las transferencias de mayor escala son los eucariotas rotíferos bdelloidea, que han recibido una serie de genes de bacterias, hongos y plantas. Los virus también se puede llevar a ADN entre organismos, permitiendo la transferencia de genes incluso a través de dominios biológicos.
La transferencia de genes a gran escala también se ha producido entre los ancestros de las células eucariotas y bacterias , durante la adquisición de cloroplastos y mitocondrias . Es posible que los propios eucariotas se originaron de transferencia horizontal de genes entre bacterias y arqueas.
Mecanismos
De un Perspectiva neo-darwiniano, la evolución se produce cuando hay cambios en las frecuencias de alelos en una población de mestizaje organismos. Por ejemplo, el alelo para el color negro en una población de polillas cada vez más común. Mecanismos que pueden conducir a cambios en las frecuencias de los alelos incluyen la selección natural , deriva genética, autostop genética, mutación y el flujo de genes.
Selección natural
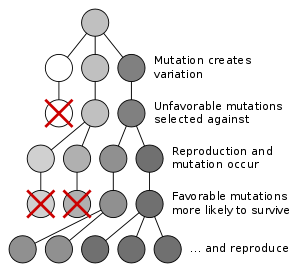
La evolución por medio de la selección natural es el proceso por el cual las mutaciones genéticas que aumentan la reproducción ser y seguir siendo más común en las sucesivas generaciones de una población. A menudo se ha llamado un mecanismo de "auto-evidente", ya que se sigue necesariamente de tres hechos simples:
- Existe variación hereditaria en las poblaciones de organismos.
- Los organismos producen más descendencia que pueden sobrevivir.
- Estas crías varían en su capacidad para sobrevivir y reproducirse.
Estas condiciones producen la competencia entre organismos para la supervivencia y la reproducción. En consecuencia, los organismos con rasgos que les dan una ventaja sobre sus competidores pasaron estos rasgos ventajosos sobre, mientras que los rasgos que no confieren una ventaja que no se transmiten a la siguiente generación.
El concepto central de la selección natural es la aptitud evolutiva de un organismo. Fitness se mide por la capacidad de un organismo para sobrevivir y reproducirse, lo que determina la magnitud de su contribución genética a la siguiente generación. Sin embargo, la aptitud no es el mismo que el número total de hijos: en lugar de la aptitud está indicado por la proporción de las generaciones posteriores que llevan genes de un organismo. Por ejemplo, si un organismo puede sobrevivir bien y reproducirse rápidamente, pero su descendencia eran demasiado pequeños y débiles para sobrevivir, este organismo tendría poco contribución genética a las generaciones futuras y que por lo tanto tienen baja aptitud.
Si un alelo aumenta la aptitud más de los otros alelos de ese gen, a continuación, con cada generación este alelo se hará más común dentro de la población. Estos rasgos se dice que están "seleccionado para". Los ejemplos de rasgos que pueden aumentar la aptitud se han mejorado la supervivencia y el aumento fecundidad. Por el contrario, la menor aptitud causado por tener un menos beneficiosos o nocivos resultados de los alelos en este alelo cada vez más raras - que se "selecciona contra". Es importante destacar que la aptitud de un alelo no es una característica fija; si el entorno cambia, previamente rasgos neutrales o perjudiciales pueden llegar a ser los rasgos beneficiosos y previamente beneficiosos se vuelven dañinos. Sin embargo, aunque la dirección de la selección hace marcha atrás de esta forma, los rasgos que se perdieron en el pasado no puede volver a evolucionar en una forma idéntica (véase La ley de Dollo).
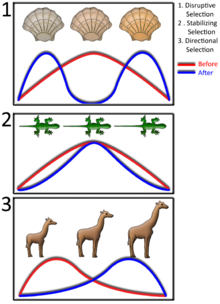
La selección natural dentro de una población de un rasgo que puede variar a través de una serie de valores, como la altura, se pueden clasificar en tres tipos diferentes. El primero es selección direccional, que es un cambio en el valor promedio de un rasgo en el tiempo - por ejemplo, organismos conseguir lentamente más alto. En segundo lugar, selección disruptiva es la selección de valores rasgo extremas ya menudo resulta en dos valores diferentes convertirse más común, con la selección contra el valor medio. Esto sería cuando los organismos, ya sea a corto o altos tenían una ventaja, pero no los de mediana estatura. Finalmente, en la estabilización de la selección no es la selección contra rasgo valores extremos en ambos extremos, lo que provoca una disminución de la varianza alrededor del valor promedio y menos diversidad. Esto, por ejemplo, causar organismos a convertirse lentamente todos la misma altura.
Un caso especial de la selección natural es la selección sexual, que es la selección para cualquier rasgo que aumenta el éxito de apareamiento al aumentar el atractivo de un organismo a parejas potenciales. Rasgos que evolucionaron a través de la selección sexual son particularmente acentuado en los machos de algunas especies animales, a pesar de rasgos tales como astas engorrosos, llamadas de apareamiento o colores brillantes que atraen a los depredadores, la disminución de la supervivencia de los hombres individuales. Esta desventaja de supervivencia se equilibra con mayor éxito reproductivo en los hombres que muestran estos difícil de falsificar, seleccionado sexualmente rasgos.
La selección natural más generalmente hace que la naturaleza de la medida contra la cual los individuos y los rasgos individuales, son más o menos probabilidades de sobrevivir. "Naturaleza" en este sentido se refiere a un ecosistema, es decir, un sistema en el que los organismos interactúan con cada otro elemento, física, así como biológica, en sus locales medio ambiente. Eugene Odum, uno de los fundadores de la ecología, define un ecosistema como: "Cualquier unidad que incluye todos los organismos ... en una zona determinada interactuando con el entorno físico para que un flujo de energía conduce a una estructura claramente definida trófica, la diversidad biótica y los ciclos de materiales (por ejemplo: intercambio de materiales entre los seres vivos y no vivos partes). dentro del sistema " Cada población dentro de un ecosistema ocupa una clara nicho, o posición, con variaciones en su relación con otras partes del sistema. Estas relaciones implican la historia de vida del organismo, su posición en el cadena alimentaria y su distribución geográfica. Esta amplia comprensión de la naturaleza permite a los científicos para delinear las fuerzas específicas que, en conjunto, conforman la selección natural.
La selección natural puede actuar en diferentes niveles de organización, como los genes, células, organismos individuales, grupos de organismos y especies. La selección puede actuar en varios niveles al mismo tiempo. Un ejemplo de selección que ocurre por debajo del nivel del organismo individual se llama genes transposones, que pueden replicarse y propagarse a través de una genoma. Selección en un nivel por encima del individuo, tales como selección de grupo, puede permitir que la evolución de la cooperación, como veremos a continuación.
Mutación Biased
Además de ser una fuente importante de variación, mutación también puede funcionar como un mecanismo de la evolución cuando hay diferentes probabilidades a nivel molecular para diferentes mutaciones que se producen, un proceso conocido como sesgo mutación. Si dos genotipos, por ejemplo uno con el nucleótido G y otro con el nucleótido A en la misma posición, tienen la misma forma física, pero la mutación de G a A sucede con más frecuencia de la mutación de la A a la G, entonces genotipos con A tenderá a evolucionar. Inserción diferente vs. sesgos mutación por deleción en diferentes taxones puede dar lugar a la evolución de los diferentes tamaños del genoma. Sesgos de desarrollo o mutacionales también se han observado en evolución morfológica. Por ejemplo, de acuerdo con la fenotipo primera teoría de la evolución, las mutaciones con el tiempo puede causar la asimilación genética de los rasgos que antes eran inducida por el medio ambiente.
Efectos de polarización mutación se superponen a otros procesos. Si la selección favorecería ya sea uno de cada dos mutaciones, pero no hay ninguna ventaja adicional de tener ambos, entonces la mutación que se produce con más frecuencia es la que es más probable que se fija en una población. Las mutaciones que conducen a la pérdida de la función de un gen son mucho más comunes que las mutaciones que producen un nuevo gen, completamente funcional. La mayor pérdida de función de las mutaciones son seleccionados en contra. Pero cuando la selección es débil, el sesgo de mutación hacia la pérdida de la función puede afectar a la evolución. Por ejemplo, los pigmentos ya no son útiles cuando los animales viven en la oscuridad de las cuevas, y tienden a perderse. Este tipo de pérdida de la función puede ocurrir debido al sesgo de la mutación, y / o porque la función tuvo un costo, y una vez que el beneficio de la función desapareció, la selección natural conduce a la pérdida. Pérdida de capacidad de esporulación en una bacteria durante la evolución de laboratorio parece haber sido causado por el sesgo de la mutación, en lugar de la selección natural contra el costo de mantenimiento de la capacidad de esporulación. Cuando no hay ninguna selección para la pérdida de la función, la velocidad a la cual la pérdida evoluciona depende más de la tasa de mutación que lo hace en el tamaño efectivo de la población, lo que indica que es impulsado más por el sesgo de la mutación que por deriva genética.
Deriva genética
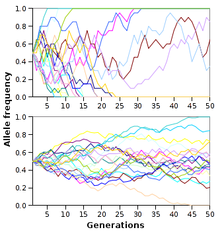
La deriva genética es el cambio en frecuencia de los alelos de una generación a la siguiente que se debe a que los alelos están sujetos a error de muestreo. Como resultado, cuando las fuerzas selectivas están ausentes o son relativamente débil, frecuencias de alelos tienden a "deriva" hacia arriba o hacia abajo al azar (en una paseo aleatorio). Esta deriva se detiene cuando un alelo se vuelve eventualmente fijo, ya sea por desaparecer de la población, o la sustitución de los otros alelos por completo. Por lo tanto, la deriva genética puede eliminar algunos alelos de una población debido a la casualidad. Incluso en ausencia de fuerzas selectivas, la deriva genética puede causar dos poblaciones separadas que comenzaron con la misma estructura genética que se separan en dos poblaciones divergentes con diferentes conjuntos de alelos.
Por lo general es difícil medir la importancia relativa de los procesos de selección y neutros, incluyendo la deriva. La importancia relativa de las fuerzas adaptativos y no adaptativos en el impulso de cambio evolutivo es un área de investigación actual .
La teoría neutral de la evolución molecular propuso que la mayoría de los cambios evolutivos son el resultado de la fijación de mutaciones neutrales por deriva genética. Por lo tanto, en este modelo, la mayoría de los cambios genéticos en una población son el resultado de la presión constante mutación y la deriva genética. Esta forma de la teoría neutral está ahora abandonada en gran medida, ya que no parece para adaptarse a la variación genética visto en la naturaleza. Sin embargo, una versión más reciente y mejor apoyado por el de este modelo es la casi teoría neutral, donde una mutación que sería neutral en una pequeña población no es necesariamente neutral en una población grande. Otras teorías alternativas proponen que la deriva genética es eclipsada por otras fuerzas estocásticos en evolución, tales como autostop genética, también conocido como proyecto genético.
El tiempo para un alelo neutral para que se fijen por la deriva genética depende del tamaño de la población, con la fijación que ocurre más rápidamente en las poblaciones más pequeñas. El número de individuos en una población no es crítica, pero en lugar de una medida conocida como la tamaño efectivo de la población. El efectivo de la población es generalmente menor que la población total ya que toma en cuenta factores como el nivel de endogamia y la etapa del ciclo de vida en el que la población es el más pequeño. El tamaño efectivo de la población puede no ser la misma para cada gen en la misma población.
Autostop Genética
La recombinación permite alelos en la misma cadena de ADN que se separan. Sin embargo, la tasa de recombinación es baja (aproximadamente dos eventos por cromosoma por generación). Como resultado, los genes cercanos juntos en un cromosoma pueden no siempre ser mezcladas de distancia el uno del otro y los genes que están muy juntas tienden a ser heredados juntos, un fenómeno conocido como vinculación. Esta tendencia se mide mediante la búsqueda de la frecuencia se producen dos alelos juntos en un solo cromosoma en comparación con expectativas, que se llama su desequilibrio de ligamiento. Un conjunto de alelos que generalmente se hereda en un grupo se denomina haplotipo. Esto puede ser importante cuando un alelo en un haplotipo particular es fuertemente beneficioso: la selección natural puede conducir una barrido selectivo que también hará que los otros alelos del haplotipo a ser más común en la población; este efecto se llama autostop genética o proyecto genético.Proyecto genética causada por el hecho de que algunos genes neutros están genéticamente relacionados con otros que se encuentran bajo la selección puede ser capturada parcialmente por un tamaño de la población eficaz apropiada.
El flujo de genes
El flujo de genes es el intercambio de genes entre poblaciones y entre las especies. La presencia o ausencia de flujo genético cambia radicalmente el curso de la evolución. Debido a la complejidad de los organismos, las dos poblaciones totalmente aisladas eventualmente evolucionar incompatibilidades genéticas a través de procesos neutros, como en el modelo de Bateson-Dobzhansky-Muller, aunque ambas poblaciones permanecen esencialmente idénticas en cuanto a su adaptación al medio.
Si la diferenciación genética entre las poblaciones se desarrolla, el flujo de genes entre poblaciones puede introducir rasgos o alelos que son desventajosos en la población local y esto puede llevar a organismo dentro de estas poblaciones de evolucionar los mecanismos que impiden que se acoplan con las poblaciones genéticamente distantes, finalmente resulta en la aparición de nuevo especies. Por lo tanto, el intercambio de información genética entre los individuos es de fundamental importancia para el desarrollo del concepto de especie biológica (BSC).
Durante el desarrollo de la síntesis moderna, Sewall Wright desarrolló su teoría del equilibrio cambiante que el flujo genético entre las poblaciones parcialmente aisladas era un aspecto importante de la evolución adaptativa. Sin embargo, recientemente ha habido críticas sustancial de la importancia de la teoría del equilibrio cambiante.
Resultados
Evolución influye en todos los aspectos de la forma y el comportamiento de los organismos. El más destacado son las de comportamiento y físicas específicas adaptaciones que son el resultado de la selección natural. Estas adaptaciones aumentan la aptitud ayudando actividades como la búsqueda de alimentos, evitar a los depredadores o atraer compañeros. Los organismos también pueden responder a la selección por cooperar entre sí, por lo general, ayudando a sus familiares o participar en mutuamente beneficiosa simbiosis. A más largo plazo, la evolución produce nuevas especies a través de las poblaciones ancestrales división de organismos en nuevos grupos que pueden o no quieren cruzarse.
Estos resultados de la evolución a veces se dividen en macroevolución, que es la evolución que se produce en o sobre el nivel de las especies, tales como la extinción y la especiación y microevolución, que es más pequeños cambios evolutivos, tales como adaptaciones, dentro de una especie o población. En general, la macroevolución es considerado como el resultado de largos períodos de microevolución. Por lo tanto, la distinción entre micro y macro-evolución no es un fundamental uno - la diferencia es simplemente el tiempo involucrado. Sin embargo, en la macroevolución, los rasgos de toda la especie pueden ser importantes. Por ejemplo, una gran cantidad de variación entre los individuos de una especie permite adaptarse rápidamente a nuevos hábitats, lo que reduce la posibilidad de que se extingan, mientras que un amplio rango geográfico aumenta la probabilidad de especiación, por lo que es más probable que una parte de la población lo hará aislarse. En este sentido, la microevolución y macroevolución podría implicar la selección en los diferentes niveles - con microevolución que actúa sobre los genes y organismos, en comparación con los procesos macroevolutivos tales como la selección de especies que actúan sobre especies enteras y que afectan a sus tasas de especiación y extinción.
Un error común es que la evolución tiene metas o planes a largo plazo; realista, sin embargo, la evolución no tiene ninguna meta a largo plazo y no produce necesariamente una mayor complejidad. Aunque especies complejas han evolucionado, que se produzcan como efecto secundario del número total de organismos cada vez mayores y las formas simples de la vida siguen siendo más común en la biosfera. Por ejemplo, la gran mayoría de las especies son microscópicos procariotas, que constituyen cerca de la mitad del mundo biomasa a pesar de su pequeño tamaño, y constituyen la gran mayoría de la biodiversidad de la Tierra. Por lo tanto, los organismos simples han sido la forma dominante de la vida en la Tierra a lo largo de su historia y siguen siendo la principal forma de vida hasta el día de hoy, con la vida compleja única apareciendo más diversa, ya que es más notable. De hecho, la evolución de los microorganismos es particularmente importante para la investigación evolutiva moderna , ya que su rápida reproducción permite el estudio de la evolución experimental y la observación de la evolución y la adaptación en tiempo real.
Adaptación
La adaptación es el proceso que hace que los organismos mejor adaptados a su hábitat. Además, la adaptación término puede referirse a un rasgo que es importante para la supervivencia de un organismo. Por ejemplo, la adaptación de los dientes de los caballos para la trituración de hierba. Al utilizar el término de adaptación para el proceso evolutivo y rasgo de adaptación para el producto (la parte o función corporal), los dos sentidos de la palabra puede ser distinguido. Las adaptaciones se producen por selección natural . Las siguientes definiciones se deben a Theodosius Dobzhansky.
- La adaptaciónes el proceso evolutivo mediante el cual un organismo se convierte en mejores condiciones para vivir en suhábitat o hábitats.
- Capacidad de adaptaciónes el estado de ser adaptada: el grado en que un organismo es capaz de vivir y reproducirse en un determinado conjunto de hábitats.
- Unrasgo de adaptaciónes un aspecto de la pauta de desarrollo del organismo que permite o aumenta la probabilidad de ese organismo sobrevivir y reproducirse.
La adaptación puede causar ya sea la ganancia de una nueva característica, o la pérdida de una característica ancestral. Un ejemplo que muestra ambos tipos de cambio es la adaptación bacteriana a los antibióticos de selección, con cambios genéticos que causan resistencia a los antibióticos por tanto la modificación de la diana del fármaco, o el aumento de la actividad de los transportadores que bombean el fármaco fuera de la célula. Otros ejemplos notables son las bacterias Escherichia coli evolución de la capacidad de utilizar ácido cítrico como un nutriente en un experimento de laboratorio a largo plazo, Flavobacterium evolucionando una nueva enzima que permite que estas bacterias crezcan en los subproductos de la fabricación de nylon, y la bacteria del suelo Sphingobium evolución una nueva vía metabólica que degrada la sintética pesticidas pentaclorofenol. Una idea interesante, pero aún controversial es que algunas adaptaciones pueden aumentar la capacidad de los organismos para generar diversidad genética y adaptación por selección natural (el aumento de los organismos ' evolvability).
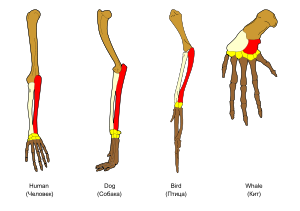
La adaptación se produce a través de la modificación gradual de las estructuras existentes. En consecuencia, las estructuras con organización interna similar pueden tener diferentes funciones en los organismos relacionados. Este es el resultado de una única estructura ancestral que está adaptado para funcionar de diferentes maneras. Los huesos dentro de murciélago alas, por ejemplo, son muy similares a aquellos en ratones pies y manos de primates, debido a la bajada de todas estas estructuras de un ancestro común de mamíferos. Sin embargo, como todos los organismos vivos están relacionados en cierta medida, incluso órganos que parecen tener poca o ninguna similitud estructural, tales como artrópodos , calamares y los ojos de vertebrados, o las extremidades y las alas de los artrópodos y vertebrados, puede depender de un conjunto común de genes homólogos que controlan su montaje y función; se llama homología profunda.
Durante la evolución, algunas estructuras pueden perder su función original y se convierten en estructuras vestigiales . Tales estructuras pueden tener poca o ninguna función en una especie de corriente, sin embargo tienen una función clara en especie ancestral, u otras especies estrechamente relacionadas. Los ejemplos incluyen pseudogenes, los restos no funcionales de los ojos de los peces ciegos cueva-vivienda, alas en aves no voladoras, y la presencia de huesos de la cadera en las ballenas y serpientes. Ejemplos de estructuras vestigiales en los seres humanos incluyen las muelas del juicio, el cóccix, el apéndice vermiforme y otros vestigios de comportamiento, como la piel de gallina y reflejos primitivos.
Sin embargo, muchos de los rasgos que parecen ser simples adaptaciones son de hecho exaptaciones: estructuras originalmente adaptadas para una función, pero que casualmente se convirtió en algo útil para alguna otra función en el proceso. Un ejemplo es el lagarto africano Holaspis guentheri , que desarrolló una cabeza extremadamente plana para esconderse en las grietas, como se puede ver mirando a sus parientes cercanos. Sin embargo, en esta especie, la cabeza se ha vuelto tan aplastada que ayuda a deslizarse de árbol en árbol, una exaptación. Dentro de las células, las máquinas moleculares como la bacteriana flagelos y maquinaria proteica clasificación evolucionaron mediante la contratación de varias proteínas pre-existente que anteriormente tenían diferentes funciones. Otro ejemplo es el reclutamiento de enzimas de la glicólisis y el metabolismo de xenobióticos para servir como proteínas estructurales llamadas cristalinas dentro de las lentes de los organismos ' ojos .
Un principio fundamental de la ecología es el de exclusión competitiva: no hay dos especies pueden ocupar el mismo nicho en el mismo ambiente por un largo tiempo. En consecuencia, la selección natural tenderá a obligar a las especies para adaptarse a diferentes nichos ecológicos. Esto puede significar que, por ejemplo, dos especies de peces cíclidos se adaptan a vivir en diferentes hábitats, lo que minimizará la competencia entre ellos por la comida.
Un área de investigación actual en biología del desarrollo evolutivo es la base del desarrollo de las adaptaciones y exaptaciones. Esta investigación aborda el origen y evolución del desarrollo embrionario y cómo las modificaciones de los procesos de desarrollo de desarrollo y producen nuevas características. Estos estudios han demostrado que la evolución puede alterar el desarrollo para producir nuevas estructuras, tales como estructuras óseas embrionarias que se desarrollan en la mandíbula en su lugar otros animales que forman parte del oído medio en los mamíferos. También es posible para las estructuras que se han perdido en la evolución a reaparecer debido a cambios en los genes de desarrollo, tales como una mutación en pollos que causan embriones para crecer dientes similares a los de cocodrilos . Ahora está cada vez más claro que la mayoría de las alteraciones en la forma de organismos son debido a cambios en un pequeño conjunto de genes conservados.
Co-evolución

Las interacciones entre los organismos pueden producir tanto los conflictos y la cooperación. Cuando la interacción es entre pares de especies, tales como un patógeno y un huésped, o un depredador y su presa, estas especies pueden desarrollar conjuntos de adaptaciones emparejados. En este caso, la evolución de una especie provoca adaptaciones en una segunda especie. Estos cambios en la segunda especie entonces, a su vez, provocan nuevas adaptaciones en la primera especie. Este ciclo de selección y la respuesta se llama co-evolución. Un ejemplo es la producción de tetrodotoxina en el tritón-áspera piel y la evolución de la resistencia a la tetrodotoxina en su depredador, la serpiente de liga común. En este par depredador-presa, una carrera de armamentos evolutiva ha producido altos niveles de la toxina en el tritón y correspondientemente altos niveles de resistencia a la toxina de la serpiente.
Cooperación
No todos los co-evolucionado interacciones entre las especies implican conflicto. Muchos casos de interacciones mutuamente beneficiosas han evolucionado. Por ejemplo, existe una cooperación extrema entre las plantas y los hongos micorrizas que crecen en sus raíces y ayudan a la planta a absorber nutrientes del suelo. Esto es un relación recíproca como las plantas proporcionan los hongos con los azúcares de la fotosíntesis. Aquí, los hongos crecen en realidad dentro de las células vegetales, lo que les permite intercambiar nutrientes con sus anfitriones, al enviar señales que suprimen la planta del sistema inmunológico .
Coaliciones entre organismos de la misma especie también han evolucionado. Un caso extremo es el eusocialidad encontrado en los insectos sociales, como las abejas , termitas y hormigas , donde se alimentan de insectos estériles y custodian el pequeño número de organismos en una colonia que son capaces de reproducirse. En una escala aún más pequeña, las células somáticas que componen el cuerpo de un animal limitan su reproducción, para mantener un organismo estable, que entonces soporta un pequeño número de animales de la células germinales para producir descendencia. En este caso, las células somáticas responden a las señales específicas que les sea crezca instruyen, permanecen como están, o mueren. Si las células ignoran estas señales y se multiplican de manera inapropiada, su crecimiento incontrolado provoca cáncer.
Esta cooperación dentro de las especies puede haber evolucionado a través del proceso de selección de parentesco, que es donde un organismo actúa para ayudar a elevar la descendencia de un familiar. Esta actividad se selecciona para porque si la ayuda individual contiene alelos que promueven la actividad ayudando, es probable que su pariente será también contener estos alelos y por lo tanto los alelos será transmitido. Otros procesos que pueden promover la cooperación incluyen la selección de grupos, donde la cooperación proporciona beneficios a un grupo de organismos.
La especiación
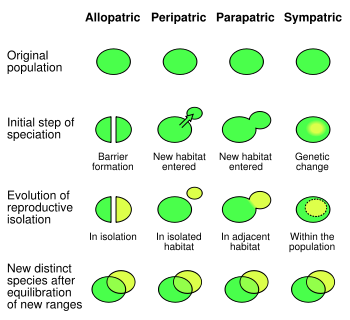
La especiación es el proceso donde una especie diverge en dos o más especies descendientes.
Hay múltiples maneras de definir el concepto de "especies". La elección de la definición depende de las particularidades de la especie en cuestión. Por ejemplo, algunos conceptos de especie se aplican más fácilmente hacia los organismos de reproducción sexual, mientras que otras se prestan mejor a los organismos asexuales. A pesar de la diversidad de los distintos conceptos de especie, estos diversos conceptos se pueden colocar en uno de los tres enfoques filosóficas generales: el entrecruzamiento, ecológica y filogenético. El concepto de especie biológica (BSC) es un ejemplo clásico del enfoque mestizaje. Definido por Ernst Mayr en 1942, el BSC afirma que "las especies son grupos de real o potencialmente cruzándose poblaciones naturales, que están reproductivamente aislados de otros grupos". A pesar de su amplio uso y de largo plazo, el BSC como los demás no está exenta de controversia, por ejemplo debido a que estos conceptos no se pueden aplicar a los procariotas, y esto se llama el problema de las especies. Algunos investigadores han intentado una definición monista unificador de las especies, mientras que otros adoptan un enfoque pluralista y sugieren que puede haber diferentes maneras de interpretar lógicamente la definición de una especie. "
Se requieren Barreras para la reproducción entre dos poblaciones divergentes sexuales para las poblaciones a convertirse en nuevas especies. El flujo de genes puede retardar este proceso mediante la difusión de las nuevas variantes genéticas también a las otras poblaciones. Dependiendo de lo lejos dos especies se han ido distanciando ya que su antepasado común más reciente, todavía puede ser posible para ellos para producir descendencia, al igual que con los caballos y los burros de apareamiento para producir mulas. Tal híbridos son generalmente infértil. En este caso, las especies estrechamente relacionadas pueden cruzarse con regularidad, pero los híbridos serán seleccionados en contra y las especies permanecerán distinta. Sin embargo, los híbridos viables son ocasionalmente formado y estas nuevas especies pueden tener ya sea propiedades intermedias entre las especies parentales, o poseer un totalmente nuevo fenotipo. La importancia de la hibridación en la producción de nuevas especies de animales no está clara, aunque los casos se han visto en muchos tipos de animales, con la rana arbórea gris es un ejemplo particularmente bien estudiado.
La especiación se ha observado varias veces tanto en condiciones controladas de laboratorio y en la naturaleza. En los organismos que se reproducen sexualmente, resultados de especiación de aislamiento reproductivo seguidas de divergencia genealógica. Hay cuatro mecanismos de especiación. El más común en los animales es la especiación alopátrica, que se produce en las poblaciones aisladas geográficamente inicialmente, como por la fragmentación del hábitat o migración. Selección en estas condiciones puede producir cambios muy rápidos en la apariencia y el comportamiento de los organismos. Como la selección y la deriva acto independientemente en las poblaciones aisladas de el resto de sus especies, la separación puede eventualmente producir organismos que no pueden cruzarse.
El segundo mecanismo de especiación es la especiación peripátrica, que ocurre cuando las pequeñas poblaciones de organismos se aíslan en un nuevo entorno. Esto difiere de la especiación alopátrica en que las poblaciones aisladas son numéricamente mucho más pequeña que la población parental. Aquí, el efecto fundador provoca especiación rápida después de un aumento de la consanguinidad aumenta selección en homocigosis, lo que lleva a un cambio genético rápido.
El tercer mecanismo de especiación es especiación parapátrica. Esto es similar a peripátrica especiación en que una pequeña población entra en un nuevo hábitat, pero difiere en que no hay ninguna separación física entre estas dos poblaciones. En lugar de ello, la especiación resulta de la evolución de los mecanismos que reducen el flujo de genes entre las dos poblaciones. Generalmente esto ocurre cuando ha habido un cambio drástico en el entorno en el hábitat de las especies parentales. Un ejemplo es la hierba odoratum Anthoxanthum , que puede someterse a la especiación parapatric en respuesta a la contaminación por metales localizada de las minas. Aquí, las plantas evolucionar que tienen resistencia a altos niveles de metales en el suelo. Selección contra el entrecruzamiento con la población parental de metal sensible produjo un cambio gradual en el tiempo de floración de las plantas de metal resistente, que finalmente producen aislamiento reproductivo completa. Selección contra híbridos entre las dos poblaciones puede causar refuerzo , que es la evolución de los rasgos que promueven el apareamiento dentro de una especie, así como el desplazamiento de caracteres, que es cuando dos especies se vuelven más clara en apariencia.
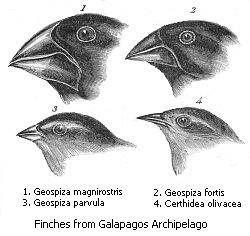
Finalmente, en especies simpátricas especiación divergen sin aislamiento geográfico o cambios en el hábitat. Esta forma es raro ya que incluso una pequeña cantidad de flujo de genes puede eliminar las diferencias genéticas entre las partes de una población. En general, la especiación simpátrica en animales requiere la evolución de ambas diferencias genéticas y el apareamiento no aleatorio, para permitir el aislamiento reproductivo de evolucionar.
Un tipo de especiación simpátrica implica el cruzamiento de dos especies relacionadas para producir una nueva especie híbrida. Esto no es común en los animales como animales híbridos son generalmente estéril. Esto es porque durante la meiosis los cromosomas homólogos de cada uno de los padres son de diferentes especies y no pueden emparejarse con éxito. Sin embargo, es más común en las plantas porque las plantas a menudo duplican su número de cromosomas, para formar poliploides. Esto permite que los cromosomas de cada especie de los padres para formar los correspondientes pares durante la meiosis, ya que los cromosomas de cada padre están representados por un par ya. Un ejemplo de un evento tan especiación es cuando las especies de plantas de Arabidopsis thaliana y Arabidopsis Arenosa cruzada criados para dar a la nueva especie Arabidopsis suecica . Esto sucedió hace unos 20.000 años, y el proceso de especiación se ha repetido en el laboratorio, lo que permite el estudio de los mecanismos genéticos implicados en este proceso. De hecho, el cromosoma duplicar dentro de una especie puede ser una causa común de aislamiento reproductivo, ya que la mitad de los cromosomas duplicados serán inigualables cuando se cría con organismos sin doblar.
Eventos de especiación son importantes en la teoría del equilibrio puntuado , que representa el patrón en el registro fósil de "ráfagas" cortos de evolución intercalados con períodos relativamente largos de estasis, donde las especies se mantienen relativamente sin cambios. En esta teoría, la especiación y evolución rápida están vinculados, con la selección natural y la deriva genética que actúa con más fuerza sobre los organismos sometidos a la especiación en nuevos hábitats o poblaciones pequeñas. Como resultado, los períodos de estancamiento en los registros fósiles corresponden a la población de los padres y los organismos sometidos a la especiación y evolución rápida se encuentran en pequeñas poblaciones o geográficamente restringidas hábitats y rara vez, por lo tanto se conservan como fósiles.
Extinción

La extinción es la desaparición de una especie entera. La extinción no es un evento inusual, como especies aparecen regularmente a través de la especiación y desaparecen a través de la extinción. Casi todas las especies animales y vegetales que han vivido en la Tierra se han extinguido, y la extinción parece ser el destino final de todas las especies. Estas extinciones han sucedido continuamente a lo largo de la historia de la vida, aunque la tasa de picos de extinción en masa ocasionales eventos de extinción. La Extinción masiva del Cretácico-Terciario, durante el cual los dinosaurios no aviares se extinguieron, es bien conocida la mayoría, pero el anterior Pérmico evento de extinción -Triassic era aún más grave, con aproximadamente el 96% de las especies conducidos a la extinción. La evento de extinción del Holoceno es una extinción masiva en curso asociada con la expansión de la humanidad en todo el mundo durante los últimos miles de años. Las tasas de extinción actuales son 100-1000 veces mayor que la tasa de fondo y hasta un 30% de las especies actuales puede ser extinguido a mediados del siglo 21. Las actividades humanas son ahora la principal causa de la extinción en curso; el calentamiento global puede acelerar aún más en el futuro.
El papel de extinción en la evolución no es muy bien entendida y puede depender de que se considera el tipo de extinción. Las causas de los eventos de extinción "de bajo nivel" continuas, que constituyen la mayoría de las extinciones, pueden ser el resultado de la competencia entre las especies por los recursos limitados ( exclusión competitiva). Si una especie puede competir fuera otro, esto podría producir la selección de especies, con las especies más en forma de sobrevivir y de las otras especies de ser conducido a la extinción. Las extinciones masivas intermitentes también son importantes, pero en lugar de actuar como una fuerza selectiva, que reducen drásticamente la diversidad de una manera no específica y promover ráfagas de rápida evolución y especiación en los supervivientes.
Historia evolutiva de la vida
Origen de la vida
Química altamente energético se cree que ha producido una molécula auto-replicante alrededor de 4000 millones años atrás, y la mitad de mil millones de años más tarde, el último ancestro común de toda la vida existió. El actual consenso científico es que la compleja bioquímica que conforma la vida vino de reacciones químicas sencillas. El comienzo de la vida puede haber incluido moléculas auto-replicantes como ARN y el montaje de las células simples.
Ascendencia común
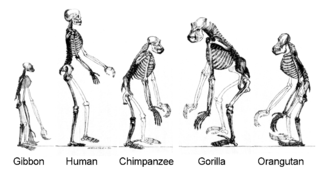
Todos los organismos en la Tierra descienden de un ancestro común o reserva genética ancestral. Especies actuales son una etapa en el proceso de la evolución, con su diversidad el producto de una larga serie de eventos de especiación y extinción. La ascendencia común de los organismos se dedujo primero de cuatro hechos simples acerca de los organismos: Primero, tienen distribuciones geográficas que no se pueden explicar por la adaptación local. En segundo lugar, la diversidad de la vida no es un conjunto de organismos completamente únicos, pero los organismos que comparten similitudes morfológicas. En tercer lugar, los rasgos vestigiales sin propósito claro asemejan rasgos ancestrales funcionales y, por último, que los organismos pueden ser clasificados usando estas similitudes en una jerarquía de grupos anidados - similar a un árbol genealógico. Sin embargo, la investigación moderna ha sugerido que, debido a la transferencia horizontal de genes, este " árbol de la vida "puede ser más complicado de lo que un árbol de ramificación sencilla ya que algunos genes se han extendido de forma independiente entre especies alejadas.
Especies anteriores también han dejado registros de su historia evolutiva. Los fósiles , junto con la anatomía comparada de los organismos actuales, constituyen las características morfológicas, o anatómica, ficha. Al comparar las anatomías de ambas especies modernas y extintas, los paleontólogos pueden inferir los linajes de esas especies. Sin embargo, este enfoque es más exitoso para los organismos que tenían partes del cuerpo duras, tales como conchas, huesos o dientes. Además, como procariotas tales como bacterias y arqueas compartir un conjunto limitado de morfologías comunes, sus fósiles no proporcionan información sobre su ascendencia.
Más recientemente, la evidencia de la descendencia común ha llegado a partir del estudio de los bioquímicos similitudes entre organismos. Por ejemplo, todas las células vivas utilizan el mismo conjunto básico de nucleótidos y aminoácidos . El desarrollo de genética molecular ha revelado el registro de la evolución que queda en los organismos ' genomas: conocer cuando las especies divergieron a través del reloj molecular producida por mutaciones. Por ejemplo, estas comparaciones de secuencias de ADN han revelado que los humanos y los chimpancés comparten el 98% de sus genomas y análisis de las pocas áreas en las que difieren ayuda a arrojar luz sobre cuándo existió el ancestro común de estas especies.
Evolución de la vida
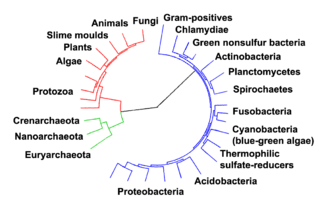
Los procariotas habitaron la Tierra de aproximadamente 3-4 millones de años atrás. No hay cambios evidentes en la morfología o la organización celular se produjeron en estos organismos en los próximos millones de años. Las células eucariotas surgieron entre 1,6 - hace 2,7 mil millones años. El siguiente cambio importante en la estructura de la célula se produjo cuando las bacterias fueron engullidos por células eucariotas, en una asociación cooperativa llamada endosimbiosis. Las bacterias envueltos y la célula huésped y luego se sometieron a la co-evolución, con la bacteria que evoluciona en cualquiera de las mitocondrias o hidrogenosomas. Otra inmersión de organismos de cianobacterias como llevó a la formación de cloroplastos en las algas y las plantas.
La historia de la vida era la de los eucariotas unicelulares, procariotas y arqueas hasta hace alrededor de 610 millones de años cuando los organismos multicelulares comenzaron a aparecer en los océanos en elperíodo Ediacara. La evolución de pluricelularidad ocurrió en múltiples eventos independientes, en organismos tan diversos como esponjas, las algas pardas , cianobacterias, moldes de limo ymixobacterias.
Poco después de la aparición de estos primeros organismos multicelulares, una cantidad notable de la diversidad biológica apareció sobre aproximadamente 10 millones de años, en un evento llamado la explosión cámbrica . Aquí, la mayoría de los tipos de animales modernos aparecieron en el registro fósil, así como linajes únicos que posteriormente se extinguieron. Se han propuesto diversos factores desencadenantes de la explosión del Cámbrico, incluyendo la acumulación de oxígeno en la atmósfera a partir de la fotosíntesis .
Hace unos 500 millones de años, las plantas y los hongos colonizaron la tierra y pronto fueron seguidos por los artrópodos y otros animales. Los insectos eran particularmente exitoso y aún hoy constituyen la mayoría de las especies animales. anfibios aparecieron por primera vez hace unos 364 millones años, seguido por los primeros amniotas y las aves hace unos 155 millones años (ambos de " reptil "-como linajes), mamíferos hace alrededor de 129 millones de años, Homininae hace unos 10 millones de años y los humanos modernos hace unos 250.000 años. Sin embargo, a pesar de la evolución de estos grandes animales, los organismos más pequeños similares a los tipos que se desarrolló a principios de este proceso siguen siendo un gran éxito y dominar la Tierra, con la mayoría de las dos de la biomasa y de las especies que son procariotas.
Aplicaciones
Conceptos y modelos utilizados en la biología evolutiva, como la selección natural, tienen muchas aplicaciones.
La selección artificial es la selección intencionada de los rasgos en una población de organismos. Esto ha sido utilizado durante miles de años en la domesticación de plantas y animales. Más recientemente, dicha selección se ha convertido en una parte vital de la ingeniería genética, con marcadores seleccionables, tales como genes de resistencia a antibióticos que se utilizan para manipular el ADN. En rondas repetidas de las proteínas de mutación y selección con propiedades valiosas han evolucionado, por ejemplo modificado enzimas y nuevos anticuerpos , en un proceso llamado evolución dirigida.
La comprensión de los cambios que se han producido durante la evolución del organismo puede revelar los genes necesarios para la construcción de partes del cuerpo, los genes que pueden estar implicados en humanos trastornos genéticos. Por ejemplo, el tetra mexicano es un albino cavefish que perdió su visión durante la evolución. Cría a diferentes poblaciones de este pez ciego producen algunos descendientes con los ojos funcionales, ya que diferentes mutaciones se habían producido en las poblaciones aisladas que se habían desarrollado en diferentes cuevas. Esto ayudó a identificar los genes necesarios para la visión y la pigmentación.
En ciencias de la computación , las simulaciones de la evolución utilizando algoritmos evolutivos y la vida artificial se inició en la década de 1960 y se extendió con la simulación de la selección artificial. evolución artificial se convirtió en un método de optimización ampliamente reconocido como resultado del trabajo de Ingo Rechenberg en la década de 1960. Utilizó estrategias de evolución a resolver problemas complejos de ingeniería. Los algoritmos genéticos, en particular, se hizo popular a través de la escritura de John Holland. Aplicaciones prácticas también incluyen la evolución automática de programas de ordenador. Los algoritmos evolutivos ahora se utilizan para resolver problemas multidimensionales más eficiente que el software producidos por diseñadores humanos y también para optimizar el diseño de sistemas.
Respuestas sociales y culturales

En el siglo 19, sobre todo después de la publicación de El Origen de las Especies en 1859, la idea de que la vida había evolucionado era una fuente activa de debate académico centrado en las implicaciones filosóficas, sociales y religiosas de la evolución. Hoy en día, la síntesis evolutiva moderna es aceptada por una gran mayoría de los científicos. Sin embargo, la evolución sigue siendo un concepto polémico para algunos teístas.
Mientras diferentes religiones y denominaciones han reconciliado sus creencias con la evolución a través de conceptos tales como la evolución teísta, hay creacionistas que creen que la evolución está en contradicción con los mitos de la creación que se encuentran en sus religiones y que crían diversas objeciones a la evolución. Como se ha demostrado por las respuestas a la publicación de Vestigios de la Historia Natural de la Creación en 1844, el aspecto más controvertido de la biología evolutiva es la implicación de la evolución humana que los seres humanos comparten un ancestro común con los simios y que las facultades mentales y morales de la humanidad tienen los mismos tipos de causas naturales como otros rasgos heredados en animales. En algunos países, especialmente los Estados Unidos, estas tensiones entre la ciencia y la religión han alimentado la actual controversia creación-evolución, un conflicto religioso centrado en la política y educacion publica. Mientras que otros campos científicos como la cosmología y las ciencias de la Tierra también conflicto con interpretaciones literales de muchos textos religiosos , las experiencias de biología evolutiva significativamente más oposición de literalistas religiosos.
La enseñanza de la evolución en las clases de biología de secundaria estadounidenses era poco común en la mayor parte de la primera mitad del siglo 20. La decisión Scopes Trial de 1925 hizo que el objeto de llegar a ser muy raro en los libros de texto de biología de secundaria estadounidenses para una generación, pero fue gradualmente re- introducido sobre una generación más tarde y legalmente protegido con el 1968 Epperson v. Arkansas decisión. Desde entonces, la creencia religiosa en competencia del creacionismo fue anulado legalmente en los planes de estudios de enseñanza secundaria en varias decisiones en los años 1970 y 1980, pero regresó en forma pseudocientífica como el diseño inteligente, ser excluido de nuevo en el 2005 Kitzmiller v. Distrito Escolar del Area de Dover caso.