


Selección natural
Sabías ...
SOS Children, que corre cerca de 200 sos escuelas en el mundo en desarrollo, organizó esta selección. Visite el sitio web de Aldeas Infantiles SOS en http://www.soschildren.org/
La selección natural es el proceso por el cual favorable heredable rasgos se vuelven más comunes en los sucesivos generaciones de una población de reproducción de los organismos , y los rasgos hereditarios desfavorables vuelto menos común, debido a la reproducción diferencial de genotipos. La selección natural actúa sobre el fenotipo, o las características observables de un organismo, de manera que los individuos con fenotipos favorables tienen más probabilidades de sobrevivir y reproducirse que los que tienen fenotipos menos favorables. El fenotipo de genética base, genotipo asociado con el fenotipo favorable, se incrementará en frecuencia en las siguientes generaciones. Con el tiempo, este proceso puede resultar en adaptaciones que se especializan organismos de concreto nichos ecológicos y eventualmente puede resultar en la aparición de nuevas especies. En otras palabras, la selección natural es el mecanismo por el cual la evolución puede tener lugar en una población de un organismo específico.
La selección natural es una de las piedras angulares de la moderna biología . El término fue introducido por Charles Darwin en su revolucionario libro de 1859 El origen de las especies en las que la selección natural fue descrita por analogía a la selección artificial, un proceso por el cual los animales con rasgos considerados deseables por los criadores humanos son favorecidos sistemáticamente para la reproducción. El concepto de la selección natural fue desarrollado originalmente en la ausencia de una teoría válida de herencia; en el momento de la escritura de Darwin, no se sabía nada de modernos genética . Aunque Gregor Mendel, el padre de la genética moderna, fue contemporáneo de Darwin, su trabajo estaría en el olvido hasta el siglo 20. La unión de tradicional La evolución darwiniana con descubrimientos posteriores en clásica y genética molecular se denomina la la síntesis evolutiva moderna. Aunque otros mecanismos de la evolución molecular, tales como la teoría neutral propuesto por Motoo Kimura, han sido identificados como causas importantes de la diversidad genética, la selección natural sigue siendo la principal explicación única para la evolución adaptativa.
Principios generales

La selección natural actúa sobre un organismo de fenotipo, o las características físicas. Fenotipo está determinado por composición genética de un organismo ( genotipo) y la entorno en el que el organismo vive. A menudo, la selección natural actúa sobre específica rasgos de un individuo, y los términos fenotipo y genotipo se utilizan estrictamente para indicar estos rasgos específicos.
Cuando diferentes organismos de una población poseen diferentes versiones de una gen para un cierto rasgo, cada una de estas versiones se conoce como una alelo. Es esta variación genética que subyace rasgos fenotípicos. Un ejemplo típico es que ciertas combinaciones de genes para color de ojos en humanos que, por ejemplo, dan lugar al fenotipo de ojos azules. (Por otro lado, cuando todos los organismos en una proporción de población el mismo alelo para un rasgo particular, y este estado es estable en el tiempo, el alelo se dice que es fijo en esa población.)
Algunos rasgos se rigen por un único gen, pero la mayoría de los rasgos están influenciadas por las interacciones de muchos genes. Una variación en uno de los muchos genes que contribuye a un rasgo puede tener sólo un pequeño efecto sobre el fenotipo; Juntos, estos genes pueden producir una serie de posibles valores fenotípicos.
Nomenclatura y el uso
El término "selección natural" tiene definiciones ligeramente diferentes en diferentes contextos. En términos simples, la "selección natural" se define con más frecuencia para operar en los rasgos hereditarios, pero a veces puede referirse a la diferencia de éxito reproductivo de fenotipos independencia de que esos fenotipos son heredables. La selección natural es "ciego" en el sentido de que el nivel de éxito reproductivo de los individuos es una función del fenotipo y no de si o en qué medida en que el fenotipo es heredable. Siguiendo uso principal de Darwin el término se utiliza a menudo para referirse tanto a la consecuencia de la selección ciego y sus mecanismos. A veces es útil distinguir explícitamente entre los mecanismos de selección y sus efectos; cuando esta distinción es importante, los científicos definen la "selección natural" específicamente como "los mecanismos que contribuyen a la selección de los individuos que se reproducen", sin tener en cuenta si la base de la selección es heredable. Esto se refiere a veces como "selección natural fenotípica.
Los rasgos que causan un mayor éxito reproductivo de un organismo se dice que ser seleccionado para, mientras que las que reducen el éxito se seleccionan en contra. La selección para un rasgo también puede dar lugar a la selección de otros rasgos correlacionados que por sí mismas no influyen directamente gimnasio. Esto puede ocurrir como resultado de pleiotropía o vinculación de genes.
Aptitud
El concepto de gimnasio es fundamental para la selección natural. Sin embargo, como con la selección natural anterior, existe una grave divergencia de opinión sobre el significado exacto del término, y Richard Dawkins logra en sus libros posteriores para evitar por completo. (Se dedica un capítulo de su libro El fenotipo extendido a discutir los diferentes sentidos en los que se utiliza el término.) A pesar de la aptitud se entiende a veces coloquialmente como una calidad que promueve la supervivencia de un individuo en particular - como se ilustra en la conocida frase supervivencia del más fuerte - la teoría evolutiva moderna define la aptitud en términos de reproducción individual. La base de este enfoque es: si un organismo vive la mitad el tiempo que otros de su especie, pero tiene el doble de hijos sobrevivientes a la edad adulta productiva, sus genes se vuelven más comunes en la población adulta de la próxima generación. Esto se conoce como la reproducción diferencial.
Aunque la selección natural actúa sobre los individuos, su efecto promedio en todos los individuos con un genotipo particular corresponde a la idoneidad de que el genotipo. Muy genotipos baja aptitud causar sus portadores tengan pocos o ningún descendiente en promedio; ejemplos incluyen muchas humana trastornos genéticos como la fibrosis quística . Condiciones como la anemia de células falciformes pueden tener baja aptitud en la población humana en general, pero debido a que confiere inmunidad frente a la malaria, que tiene un alto valor de fitness en las poblaciones que tienen altas tasas de infección de malaria. En términos generales, la aptitud de un organismo es una función de fitnesses sus alelos. Desde la aptitud es una cantidad promedio, sin embargo, es posible una mutación favorable puede surgir en un individuo que no sobreviven a la edad adulta, por razones no relacionadas.
Tipos de selección
La selección natural puede actuar en cualquier rasgo fenotípico, y la presión selectiva puede ser producido por cualquier aspecto del medio ambiente, incluyendo compañeros y congéneres, o miembros de la misma especie. Sin embargo, esto no implica que la selección natural es siempre direccional y los resultados en la evolución de adaptación; la selección natural a menudo resulta en el mantenimiento del statu quo mediante la eliminación de las variantes menos aptos.
La unidad de selección puede ser el individuo o puede ser otro nivel dentro de la jerarquía de la organización biológica, tales como genes, células, y grupos de parentesco. Todavía se debate si la selección natural actúa a nivel de grupos o especies para producir adaptaciones que benefician a un grupo más grande, no parientes. Selección en un nivel diferente tal como el gen puede resultar en un aumento de la aptitud para ese gen, mientras que al mismo tiempo la reducción de la aptitud de los individuos portadores de ese gen, en un proceso llamado conflicto intragenomic. En general, el efecto combinado de todas las presiones de selección en varios niveles determina el estado físico general de un individuo, y por lo tanto el resultado de la selección natural.
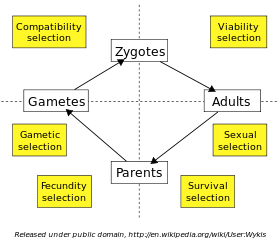
La selección natural se produce en cada etapa de la vida de un individuo. Un organismo individual debe sobrevivir hasta la edad adulta antes de que pueda reproducirse, y la selección de los que alcanzan esta etapa se llama selección viabilidad. En muchas especies, los adultos deben competir entre sí por los compañeros a través de la selección sexual, y el éxito en esta competición determina quién será el padre de la próxima generación. Cuando los individuos pueden reproducir más de una vez, una mayor supervivencia en la fase reproductiva aumenta el número de hijos, llamado selección supervivencia. La fecundidad de las hembras y los machos (por ejemplo, el gigante espermatozoides en ciertas especies de Drosophila) se puede limitar a través de la selección fecundidad. La viabilidad de producido gametos pueden diferir, mientras conflictos intragenomic como unidad meiótica entre el gametos haploides pueden dar lugar a la selección gametic o génica. Por último, la unión de algunas combinaciones de huevos y esperma podría ser más compatible que otros; esto se denomina selección de compatibilidad.
La selección sexual
También es útil hacer una distinción entre mecanicista selección ecológica y el término específico la selección sexual. selección ecológica cubre cualquier mecanismo de la selección como resultado del medio ambiente (incluyendo, por ejemplo, parientes la selección de parentesco, y sus congéneres, por ejemplo, competencia, el infanticidio), mientras que la selección sexual se refiere específicamente a la competencia entre los congéneres de los ayudantes. La selección sexual puede ser intrasexual, como en los casos de competencia entre individuos del mismo sexo en una población, o intersexual, como en los casos en que uno de los sexos controles de acceso reproductiva eligiendo entre una población de compañeros disponibles. Más comúnmente, la selección intrasexual implica la competencia entre machos y selección intersexual implica la elección femenina de machos adecuados, debido a la generalmente mayor inversión de recursos para una mujer que un hombre en un solo organismo descendencia. Sin embargo, algunas especies exhiben roles sexuales comportamiento invertido en el que son los hombres los que son más selectivos en la elección de pareja; los ejemplos más conocidos de este patrón se producen en algunos peces de la familia Syngnathidae, aunque también se han encontrado ejemplos en los probables anfibios y aves especies. Algunas de las características que se limitan a un solo sexo de una especie en particular pueden ser explicados por la selección ejercida por el otro sexo en la elección de un compañero, por ejemplo, el plumaje extravagante de algunos machos. Del mismo modo, la agresión entre miembros del mismo sexo se asocia a veces con características muy particulares, como la cornamenta de ciervos, que se utilizan en combate con otros ciervos. Más en general, la selección intrasexual se asocia a menudo con dimorfismo sexual, incluidas las diferencias en el tamaño corporal entre machos y hembras de una especie.
Un ejemplo: resistencia a los antibióticos
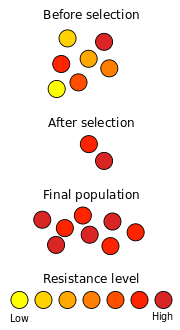
Un ejemplo bien conocido de la selección natural en acción es el desarrollo de resistencia a los antibióticos en microorganismos. Desde el descubrimiento de penicilina en 1928 por Alexander Fleming , los antibióticos se han utilizado para combatir bacterias enfermedades. Las poblaciones naturales de bacterias contienen, entre un gran número de sus miembros individuales, una variación considerable en su material genético, principalmente como resultado de mutaciones. Cuando se expone a los antibióticos, la mayoría de las bacterias mueren rápidamente, pero algunos pueden tener mutaciones que les hacen poco menos susceptible. Si la exposición a los antibióticos es corto, estos individuos sobreviven al tratamiento. Esta eliminación selectiva de los individuos inadaptados de una población es la selección natural.
Estas bacterias sobreviven entonces reproducir de nuevo, la producción de la próxima generación. Debido a la eliminación de los individuos mal adaptados en la última generación, esta población contiene más bacterias que tienen una cierta resistencia contra el antibiótico. Al mismo tiempo, se producen nuevas mutaciones, contribuyendo nueva variación genética a la variación genética existente. Las mutaciones espontáneas son muy raros, y las mutaciones ventajosas son aún más raros. Sin embargo, las poblaciones de bacterias son lo suficientemente grande que unos pocos individuos tendrán mutaciones beneficiosas. Si una nueva mutación reduce su susceptibilidad a un antibiótico, estos individuos son más probabilidades de sobrevivir cuando al lado enfrentado a ese antibiótico. Dado el tiempo suficiente, y la exposición repetida a los antibióticos, una población de bacterias resistentes a los antibióticos surgirá.
El uso generalizado y mal uso de los antibióticos ha dado lugar a una mayor resistencia microbiana a los antibióticos de uso clínico, hasta el punto de que el Staphylococcus aureus resistente a la meticilina (MRSA) se ha descrito como un " superbacteria ", debido a la amenaza que representa para la salud y su invulnerabilidad relativa a los medicamentos existentes. Las estrategias de respuesta incluyen típicamente el uso de diferentes antibióticos, más fuertes; Sin embargo, la nueva cepas de MRSA han surgido recientemente que son resistentes incluso a estos fármacos. Este es un ejemplo de lo que se conoce como una carrera de armamentos evolutiva, en la que las bacterias continúan desarrollando cepas que son menos susceptibles a los antibióticos, mientras que los investigadores médicos continúan desarrollando nuevos antibióticos que pueden matar a ellos. Una situación similar ocurre con resistencia a los pesticidas en plantas e insectos. Las carreras armamentistas no necesariamente están inducidos por el hombre; un ejemplo bien documentado implica la elaboración de la Vía de interferencia de ARN en las plantas como medio de la inmunidad innata contra los virus .
La teoría genética de la selección natural
La selección natural por sí mismo es un concepto simple, en la que las diferencias de aptitud entre fenotipos juegan un papel crucial. Es la unión de la selección natural como mecanismo con el material genético como sustrato que ofrece la mayoría de poder explicativo de la teoría
La direccionalidad de la selección
Cuando algún componente de un rasgo es heredable, la selección se altera las frecuencias de los distintos alelos o variantes del gen que produce las variantes del rasgo. La selección puede ser dividido en tres clases, sobre la base de su efecto en las frecuencias de alelos.
Selección direccional se produce cuando un alelo tiene una cierta aptitud mayor que otros, lo que resulta en un aumento en la frecuencia de ese alelo. Este proceso puede continuar hasta que el alelo es fijo y toda la población comparte el fenotipo en forma. Es la selección direccional que se ilustra en el ejemplo resistencia a los antibióticos anteriormente .
Mucho más común es la estabilización de selección (también conocido como purificador de selección), lo que disminuye la frecuencia de alelos que tienen un efecto perjudicial sobre el fenotipo - es decir, producir organismos de menor aptitud. Este proceso puede continuar hasta que el alelo se elimina de la población. Resultados purificación de selección en las características genéticas funcionales, tales como genes que codifican proteínas o secuencias reguladoras, siendo conservado con el tiempo debido a la presión selectiva contra variantes deletéreas.
Finalmente, un número de formas de equilibrio de la selección existir, que no den lugar a la fijación, pero mantener un alelo a frecuencias intermedias en una población. Esto puede ocurrir en especies diploides (es decir, aquellos que tienen dos pares de cromosomas) cuando individuos heterocigotos, que tienen diferentes alelos de cada cromosoma en una sola locus genético, tienen una mayor aptitud que individuos homocigotos que tienen dos de los mismos alelos. Se llama ventaja o overdominance heterocigoto, de los cuales el ejemplo más conocido es el de la malaria resistencia observada en los seres humanos heterocigotos que llevan sólo una copia del gen de la anemia de células falciformes . Mantenimiento de la variación alélica también puede ocurrir a través selección disruptiva o la diversificación, lo que favorece genotipos que se apartan de la media en cualquier dirección (es decir, lo contrario de overdominance), y puede resultar en una distribución bimodal de valores de los caracteres. Por último, la selección de equilibrio puede ocurrir a través de dependiente de la frecuencia de selección, donde la aptitud de un fenotipo particular depende de la distribución de otros fenotipos en la población. Los principios de la teoría de juegos se han aplicado a entender las distribuciones de fitness en estas situaciones, en particular en el estudio de la selección de parentesco y la evolución de altruismo recíproco.
Selección y la variación genética
Una porción de todas variación genética es funcionalmente neutral en que no produce ningún efecto fenotípico o diferencia significativa en la aptitud; la hipótesis de que las cuentas de esta variación de una gran fracción de la observada la diversidad genética se conoce como el teoría neutral de la evolución molecular y fue originado por Motoo Kimura. Variación neutral vez se pensó para abarcar la mayor parte de la variación genética en ADN no codificante, que se planteó la hipótesis de que se compone de " ADN basura ". Sin embargo, más recientemente, el papel funcional de ADN no codificante, tales como las funciones de regulación y de desarrollo de ARN productos genéticos, se ha estudiado en profundidad; gran parte de las secuencias de ADN no codificantes de proteínas son muy conservadas bajo una fuerte selección purificadora y por lo tanto no varían mucho de persona a persona, lo que indica que las mutaciones en estas regiones tienen consecuencias deletéreas. Cuando la variación genética no da lugar a diferencias en la aptitud, la selección no puede afectar directamente a la frecuencia de tal variación. Como resultado, la variación genética en esos sitios será más alto que en los sitios donde la variación influye la condición física.
Selección equilibrio mutación
La selección natural da como resultado la reducción de la variación genética a través de la eliminación de los individuos mal adaptados y en consecuencia de las mutaciones que causan la mala adaptación. Al mismo tiempo, se producen nuevas mutaciones, resultando en una equilibrio mutación-selección. El resultado exacto de los dos procesos depende tanto de la velocidad a la que se producen nuevas mutaciones y de la fuerza de la selección natural, que es una función de la forma desfavorable la mutación resulta ser. En consecuencia, los cambios en la tasa de mutación o de la presión de selección dará como resultado un equilibrio mutación-selección diferente.
Ligamiento genético
Ligamiento genético se produce cuando el loci de dos alelos están vinculadas, o en estrecha proximidad entre sí en el cromosoma. Durante la formación de gametos, recombinación de los resultados materiales genéticos en reorganización de los alelos. Sin embargo, la probabilidad de que una remodelación tales se produce entre dos alelos depende de la distancia entre dichos alelos; el cerrador de los alelos son el uno al otro, menos probable es que una reestructuración tal ocurrirá. En consecuencia, cuando la selección se dirige a un alelo, esto se traduce automáticamente en la selección del otro alelo también; a través de este mecanismo, la selección puede tener una fuerte influencia en los patrones de variación en el genoma.
Barridos selectivos se producen cuando un alelo se vuelve más común en una población como resultado de la selección positiva. A medida que la prevalencia de uno de los alelos aumenta, alelos ligados también se vuelven más comunes, ya sean neutros o incluso ligeramente deletéreo. Se llama autostop genética. Un fuerte resultados barrido selectivo en una región del genoma donde el seleccionado positivamente haplotipo (el alelo y sus vecinos) son esencialmente los únicos que existen en la población.
Si ha habido un barrido selectivo o no puede ser investigado por la medición desequilibrio de ligamiento, o si un determinado haplotipo está sobrerrepresentado en la población. Normalmente, resultados de recombinación genética en una reorganización de los diferentes alelos de un haplotipo, y ninguno de los haplotipos dominarán la población. Sin embargo, durante un barrido selectivo, la selección para un alelo específico también resultar en la selección de alelos vecinos. Por lo tanto, la presencia de fuerte desequilibrio de ligamiento podría indicar que se ha producido un barrido selectivo 'reciente', y esto puede ser usado para identificar sitios recientemente bajo selección.
Selección de fondo es lo contrario de un barrido selectivo. Si un sitio específico experimenta purificación de selección fuerte y persistente, la variación ligada tenderá a ser eliminados junto con él, produciendo una región en el genoma de baja variabilidad global. Debido a la selección de fondo es el resultado de nuevas mutaciones deletéreas, que pueden ocurrir al azar en cualquier haplotipo, no produce desequilibrio de ligamiento.
La evolución por medio de la selección natural
Un requisito previo para la selección natural para dar lugar a evolución adaptativa, rasgos nuevos y especiación, es la presencia de heredable variación genética que da lugar a diferencias de aptitud. La variación genética es el resultado de mutaciones, recombinaciones y alteraciones en el cariotipo (el número, forma, tamaño y disposición interna de la cromosomas). Cualquiera de estos cambios podrían tener un efecto que es altamente ventajoso o muy desfavorable, pero los efectos grandes son muy raros. En el pasado, la mayoría de los cambios en el material genético se consideraron neutro o casi neutro porque ocurrieron en ADN no codificador o resultado en una sinónimo de sustitución. Sin embargo, la investigación reciente sugiere que muchas mutaciones en el ADN no codificante tienen efectos deletéreos leves. Aunque tanto las tasas de mutación y efectos promedio de aptitud de las mutaciones son dependientes del organismo, las estimaciones a partir de datos en seres humanos han encontrado que la mayoría de las mutaciones son ligeramente deletéreo.

Por la definición de la aptitud, los individuos con mayor aptitud tienen más probabilidades de contribuir descendientes a la siguiente generación, mientras que los individuos con menor aptitud son más propensos a morir antes o dejan de reproducirse. Como resultado, los alelos que el resultado promedio en mayor aptitud se vuelven más abundantes en la próxima generación, mientras que los alelos que generalmente reducen la aptitud vuelven más raras. Si las fuerzas de selección siguen siendo los mismos para muchas generaciones, los alelos beneficiosos son cada vez más abundantes, hasta que dominan la población, mientras que los alelos con una aptitud menor desaparecen. En cada generación, nuevas mutaciones y recombinaciones surgen espontáneamente, produciendo un nuevo espectro de fenotipos. Por lo tanto, cada nueva generación se verá enriquecida por la creciente abundancia de alelos que contribuyen a los rasgos que fueron favorecidos por la selección, la mejora de estas características a lo largo de sucesivas generaciones.
Algunas mutaciones se producen en la llamada genes reguladores. Los cambios en estos pueden tener grandes efectos sobre el fenotipo del individuo porque regulan la función de muchos otros genes. La mayoría, pero no todas, las mutaciones en genes reguladores resultan en no viable cigotos. Ejemplos de mutaciones reguladoras no letales ocurren en HOX genes en los seres humanos, lo cual puede resultar en una costilla cervical o polidactilia, un aumento en el número de dedos o dedos de los pies. Cuando tales mutaciones dan como resultado una mayor aptitud, la selección natural favorecerá estos fenotipos y la novela rasgo se extenderá en la población.
Rasgos de la empresa no son inmutables; rasgos que tienen alta aptitud en un contexto ambiental pueden ser mucho menos aptos, si las condiciones ambientales cambian. En ausencia de la selección natural para preservar un rasgo tal, será más variable y deteriorarse con el tiempo, resultando posiblemente en un vestigial manifestación del rasgo. En muchas circunstancias, la estructura aparentemente vestigial puede retener una funcionalidad limitada, o puede ser co-optado por otros rasgos ventajosos en un fenómeno conocido como preadaptacion. Un ejemplo famoso de una estructura vestigial, el ojo de la rata topo ciego, se cree que para mantener la función en percepción fotoperiodo.
La especiación
La especiación requiere el apareamiento selectivo, que se traducen en un menor el flujo de genes. Apareamiento selectivo puede ser el resultado de, por ejemplo, un cambio en el entorno físico (aislamiento físico por una barrera extrínseca), o por la selección sexual resultando en apareamiento selectivo. Con el tiempo, estos subgrupos pueden divergir radicalmente para convertirse en especies diferentes, ya sea debido a las diferencias en las presiones de selección en los diferentes subgrupos, o debido a que diferentes mutaciones surgen espontáneamente en las diferentes poblaciones, o debido efectos fundadores - algunos alelos potencialmente beneficiosos pueden, por casualidad, estar presentes en sólo uno u otro de dos subgrupos de la primera vez que se separan. Un mecanismo menos conocido de la especiación se produce a través hibridación, bien documentado en las plantas y en ocasiones observa en grupos ricos en especies de animales como peces cíclidos. Tales mecanismos de especiación rápida pueden reflejar un mecanismo de cambio evolutivo conocido como equilibrio puntuado , lo que sugiere que el cambio evolutivo y en particular la especiación típicamente ocurre rápidamente después de interrumpir largos períodos de estasis.
Los cambios genéticos dentro de los grupos resultan en el incremento de incompatibilidad entre los genomas de los dos subgrupos, reduciendo así el flujo de genes entre los grupos. El flujo de genes cesará efectivamente cuando las mutaciones distintivas que caracterizan a cada subgrupo se convierten en fijos. Como sólo dos mutaciones pueden dar lugar a la especiación: si cada mutación tiene un efecto neutro o positivo sobre la aptitud cuando se producen por separado, pero un efecto negativo cuando se producen juntos, entonces la fijación de estos genes en los respectivos subgrupos dará lugar a dos reproductivamente aislados poblaciones. De acuerdo con el concepto de especie biológica, estos serán dos especies diferentes.
Desarrollo histórico

Teorías predarwinianos
Varios filósofos antiguos expresaron la idea de que la naturaleza produce una gran variedad de criaturas, aparentemente al azar, y que sólo las criaturas sobreviven que logran mantenerse a sí mismos y reproducirse con éxito; ejemplos bien conocidos incluyen Empédocles y su sucesor intelectual, Lucrecio, mientras que las ideas relacionadas fueron posteriormente refinada por Aristóteles . La lucha por la vida fue descrito más tarde por al-Jahiz en el siglo noveno. Tales argumentos clásicos fueron reintroducidos en el siglo 18 por Pierre Louis Maupertuis y otros, incluyendo a Charles Darwin abuelo 's Erasmus Darwin. Si bien estos precursores tuvieron una influencia en Darwinismo, que más tarde tuvo poca influencia en la trayectoria del pensamiento evolucionista de Charles Darwin.
Hasta principios del siglo 19, el vista en las sociedades occidentales predominante era que las diferencias entre los individuos de una especie eran poco interesantes salidas de su Ideal platónico (o typus) de clases creadas. Sin embargo, la teoría de la uniformismo en geología promovió la idea de que las fuerzas simples y débiles podrían actuar de forma continua durante largos períodos de tiempo para producir cambios radicales en el paisaje de la Tierra. El éxito de esta teoría creó conciencia de la vasta escala del tiempo geológico y hace verosímil la idea de que pequeños cambios, prácticamente imperceptibles en generaciones sucesivas podrían producir consecuencias en la escala de las diferencias entre las especies. A principios del siglo 19 evolucionistas tales como Jean Baptiste Lamarck sugirió la la herencia de los caracteres adquiridos como un mecanismo para el cambio evolutivo; rasgos adaptativos adquiridos por un organismo durante su vida podrían ser heredados por la progenie de ese organismo, causando con el tiempo transmutación de las especies. Esta teoría ha llegado a ser conocido como Lamarckismo y fue una influencia en las ideas anti-genéticos de la Estalinista soviético biólogo Trofim Lysenko.
La hipótesis de Darwin
Entre 1842 y 1844, Charles Darwin expuso su teoría de la evolución por selección natural como explicación para la adaptación y la especiación. Definió la selección natural como el "principio por el cual cada pequeña variación [de un rasgo], si es útil, se conserva". El concepto era simple pero potente: los individuos mejor adaptados a su entorno tienen más probabilidades de sobrevivir y reproducirse. Mientras haya alguna variación entre ellos, habrá un inevitable selección de individuos con las variaciones más ventajosas. Si se heredan las variaciones, entonces el éxito reproductivo diferencial dará lugar a una evolución progresiva de determinadas poblaciones de una especie, y de las poblaciones que evolucionan a ser lo suficientemente diferente eventualmente podría convertirse en especies diferentes.
Las ideas de Darwin fueron inspirados por las observaciones que había hecho sobre el viaje del Beagle, y por el trabajo de dos economistas. El primero fue Thomas Malthus , que en Ensayo sobre el principio de la población, señaló que la población (si no se controla) aumenta exponencialmente mientras que el suministro de alimentos crece sólo aritméticamente; por lo tanto, inevitables limitaciones de recursos tendrían implicaciones demográficas, lo que lleva a una "lucha por la existencia", en el que sólo los más aptos sobrevivirán. El segundo fue Adam Smith quien, en La riqueza de las naciones, identificó un mecanismo de regulación en los mercados libres, que se refirió como la " mano invisible ", lo que sugiere que los precios que se ajuste automáticamente de acuerdo con los suministros y la demanda. Así, para Darwin, el desastre que se suponía iba a ocurrir de acuerdo con Malthus se mantiene bajo control y constantemente mejorada por la competencia (o ley de la selección).
Una vez que la teoría se había formulado, Darwin era meticuloso con la recopilación y pruebas de refinación, compartiendo sus ideas sólo con unos pocos amigos; que se inspiró para publicar después del naturalista Alfred Russel Wallace concebido independientemente del principio y lo describió en un ensayo que envió a Darwin. Un arreglo fue hecho (sin el conocimiento de Wallace) para presentar su ensayo y dos escritos inéditos cortos de Darwin a la Linnean Sociedad anunciando co-descubrimiento del principio de julio 1858; Darwin publicó un informe más detallado de sus pruebas y conclusiones en El origen de las especies en 1859. En la sexta edición de El origen de las especies Darwin reconoció que otros - especialmente William Charles Wells en 1813, y Patrick Matthew en 1831 - había propuesto teorías similares, pero no los había presentado en todo o en publicaciones científicas notables.
Darwin pensaba de la selección natural por analogía a cómo los agricultores seleccionan los cultivos o el ganado de cría, a la que llamó selección artificial; en sus primeros manuscritos se refirió a una «naturaleza» que haría la selección. En ese momento, otros mecanismos de la evolución, como evolución por deriva genética todavía no se formularon de manera explícita, y Darwin se dio cuenta de que la selección era probable que sólo parte de la historia: "Estoy convencido de que [lo] ha sido el principal, pero no único medio de modificación." Para Darwin y sus contemporáneos, la selección natural era, pues, esencialmente sinónimo de evolución por selección natural . Después de la publicación de El origen de las especies, las personas educadas generalmente aceptado que la evolución había ocurrido en alguna forma. Sin embargo, la selección natural se mantuvo controversial como un mecanismo, en parte porque se percibe como demasiado débil para explicar la gama de características observadas de los organismos vivos, y en parte porque incluso los partidarios de la evolución se resistió a su 'sin guía' y el carácter no progresivo, un la respuesta que se ha caracterizado como el impedimento más importante para la aceptación de la idea. Sin embargo, algunos pensadores adoptaron con entusiasmo la selección natural; después de leer Darwin, Herbert Spencer introdujo el término supervivencia del más fuerte , que se convirtió en un resumen popular de la teoría. Aunque la frase todavía se utiliza a menudo por los no biólogos, biólogos modernos evitan porque es tautológico si más apto se lee en el sentido funcionalmente superior y se aplica a las personas en lugar de considerarse como una cantidad promediada sobre las poblaciones. En una carta a Charles Lyell en 1860 de septiembre de Darwin lamenta el uso del término 'selección natural', prefiriendo el término "natural Conservación '.
Síntesis evolutiva moderna
Sólo después de la integración de la teoría de la evolución con una apreciación estadística compleja de monje austríaco leyes de Gregor Mendel 're-descubierto "de la herencia qué la selección natural se convierten generalmente aceptada por los científicos. El trabajo de Ronald Fisher (que desarrolló el lenguaje de las matemáticas y de la selección natural en términos de los procesos genéticos subyacentes), JBS Haldane (que introdujo el concepto de "costo" de la selección natural), Sewall Wright (quien dilucidó la naturaleza de la selección y adaptación), Theodosius Dobzhansky (que estableció la idea de que la mutación, mediante la creación genética diversidad, suministrado la materia prima para la selección natural), William Hamilton (quien concibió la selección de parentesco), Ernst Mayr (que reconoció la importancia clave de aislamiento reproductivo para la especiación) y muchos otros formaron la síntesis evolutiva moderna. Esta síntesis cimentó la selección natural como fundamento de la teoría de la evolución, donde permanece en la actualidad.
Impacto de la idea
Darwin 's las ideas, junto con las de Adam Smith y Karl Marx , tuvieron una profunda influencia en el pensamiento del siglo 19. Tal vez la afirmación más radical de la teoría de la evolución por selección natural es que las "formas elaboradamente construidas, tan diferentes unos de otros, y dependientes unos de otros de manera tan compleja" evolucionaron a partir de las formas más simples de la vida por unos pocos principios simples. Esta afirmación inspiró algunos de los más fervientes partidarios de Darwin-y provocó la más profunda oposición. El radicalismo de la selección natural, de acuerdo con Stephen Jay Gould , estaba en su poder para "destronar a algunas de las comodidades más profundas y de mayor tradición del pensamiento occidental". En particular, desafió las creencias de larga data en tales conceptos como un lugar especial y sublime para los seres humanos en el mundo natural y un creador benevolente cuyas intenciones se reflejaron en orden y diseño de la naturaleza.
La teoría social y psicológica
Las implicaciones sociales de la teoría de la evolución por selección natural también se convirtió en la fuente de continua controversia. Friedrich Engels , un alemán filósofo político y co-creador de la ideología del comunismo , escribió en 1872 que "Darwin no sabía lo que una sátira amarga que escribió sobre la humanidad cuando mostró que la libre competencia, la lucha por la existencia, que los economistas celebran como el mayor logro histórico, es el estado normal del reino animal ". Interpretación de la selección natural como necesariamente "progresista", lo que lleva al aumento de 'Jugada' de la inteligencia y de la civilización, se utilizó como justificación para el colonialismo y las políticas de la eugenesia, así como las posiciones sociopolíticas más amplias ahora descrito como el darwinismo social. Konrad Lorenz ganó el Nobel Premio de Fisiología o Medicina en 1973 por su análisis del comportamiento animal en términos del papel de la selección natural (en particular la selección de grupo). Sin embargo, en Alemania en 1940, en los escritos que posteriormente repudiado, utilizó la teoría como justificación de las políticas del nazi estado. No escribió "... la selección de la dureza, el heroísmo y la utilidad social ... debe ser realizada por una institución humana, si la humanidad, a falta de factores selectivos, está a ser arruinado por la degeneración inducida domesticación. La idea racial como la base de nuestro estado ya se ha logrado mucho en este sentido ". Otros han desarrollado ideas que las sociedades y la cultura humanas evolucionan por mecanismos que son análogos a los que se aplican a la evolución de las especies.
Más recientemente, el trabajo entre los antropólogos y psicólogos ha llevado al desarrollo de la sociobiología y más tarde la psicología evolutiva, un campo que trata de explicar las características de la psicología humana en términos de adaptación al medio ambiente ancestral. El más prominente de estos ejemplos, especialmente avanzado en los primeros trabajos de Noam Chomsky y más tarde por Steven Pinker, es la hipótesis de que el cerebro humano se adapta a adquirir las reglas gramaticales de la lengua natural. Otros aspectos de la conducta humana y las estructuras sociales, de las normas culturales específicas, tales como la evitación del incesto a los patrones más amplios, como los roles de género, se han planteado la hipótesis de tener orígenes similares a las adaptaciones al ambiente temprano en el que los humanos modernos evolucionaron. Por analogía con la acción de la selección natural en los genes, el concepto de memes - "unidades de transmisión cultural", o sus equivalentes de la cultura de los genes sometidos a selección y recombinación - ha surgido, descrita por primera vez en esta forma por Richard Dawkins y posteriormente ampliado a los filósofos como Daniel Dennett como explicaciones para actividades culturales complejas, incluyendo humanos conciencia. Extensiones de la teoría de la selección natural a una amplia gama de fenómenos culturales, han sido claramente controvertido y no son ampliamente aceptados.
Información y la teoría de sistemas
En 1922, Alfred Lotka propone que la selección natural puede ser entendido como un principio físico que podría ser cuantificado con energía, un concepto que se desarrolló más tarde por Howard Odum como el principio de máxima potencia mediante el cual los sistemas evolutivos con ventaja selectiva maximizar la tasa de transformación de la energía útil. Tales conceptos son a veces relevantes en el estudio de las aplicadas termodinámica .
Los principios de la selección natural han inspirado una variedad de técnicas computacionales, tales como "suave" vida artificial, que simulan los procesos selectivos y pueden ser altamente eficientes en la 'adaptación' entidades a un entorno definido por una especifica función de aptitud. Por ejemplo, una clase de heurísticas de optimización de algoritmos conocidos como algoritmos genéticos, por primera vez por John Holland en los años 1970 y ampliadas sobre por David Goldberg, identificar soluciones óptimas de reproducción simulada y la mutación de una población de soluciones definidas por una inicial distribución de probabilidad . Tales algoritmos son particularmente útiles cuando se aplican a problemas cuya paisaje solución es muy áspero o tiene muchos mínimos locales.