

Fotosíntesis
Acerca de este escuelas selección Wikipedia
SOS ofrecen una descarga completa de esta selección de escuelas para su uso en escuelas intranets. Antes de decidir sobre el patrocinio de un niño, ¿por qué no aprender sobre diferentes obras de caridad de patrocinio primero ?
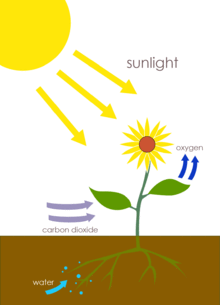
La fotosíntesis es un proceso por el cual la mayoría de las plantas y algunos otros organismos capturan la energía luminosa de la luz solar para conducir procesos químicos necesarios para alimentar los organismos ' metabolismo. En su mayor parte se combinan dióxido de carbono y agua para formar hidratos de carbono , tales como azúcares. Esto da lugar al nombre de la fotosíntesis, del griego [foto-], "luz", φώτο- y σύνθεσις [síntesis], "armar". El proceso también libera libre de oxígeno , sobre todo como un producto de desecho. Cianobacterias, la mayoría de las plantas , y la mayoría de las algas realizan la fotosíntesis, y tales organismos se llaman Fotoautótrofos. La fotosíntesis mantiene los niveles de oxígeno atmosférico y, o bien proporciona la energía necesaria para la mayoría de la vida en la Tierra, o los precursores que los organismos pueden metabolizar para crear otros compuestos compuestos del carbono. Las principales excepciones son quimiotrofos, organismos que obtienen energía a través de reacciones químicas que no son impulsados por energía de la luz.
Introducción
Aunque diferentes especies realizan la fotosíntesis de diferentes maneras, el proceso siempre comienza cuando la energía de la luz es absorbida por las estructuras de proteínas moléculas y pigmentos. Estos son los llamados centros de reacción ; que contienen diversos pigmentos, principalmente verde clorofilas. En las plantas de estas proteínas se llevan a cabo en el interior orgánulos llamados cloroplastos, principalmente en células de las hojas. Pigmentos fotosintéticos en bacterias están incrustadas en el membrana de plasma.
En estas reacciones dependientes de la luz algunas de las tiras de energía capturada electrones de sustancias adecuadas, normalmente agua, que separan los átomos de oxígeno a partir de sus iones y electrones de hidrógeno unidos. Los átomos de oxígeno se combinan en moléculas de oxígeno y son en gran medida descartados, que escapen a la atmósfera. Los electrones y los iones de hidrógeno se transfieren a un compuesto llamado fosfato de nicotinamida adenina dinucleótido (NADP +), reduciéndolo a NADPH. La mayor parte de la NADPH se consume en la generación de trifosfato de adenosina (ATP), la "moneda de energía" de las células.
En las plantas, algas y cianobacterias, azúcares se producen a partir de los productos inmediatos de la fotosíntesis por una secuencia de reacciones independientes de la luz llamado Ciclo de Calvin. Algunas otras bacterias utilizan diferentes procesos metabólicos, tales como la revertir el ciclo de Krebs. En el ciclo de Calvin, el dióxido de carbono atmosférico es incorporado en compuestos orgánicos de carbono ya existentes, tales como ribulosa bifosfato (RuBP). Esta incorporación inicial de carbono en compuestos orgánicos se denomina fijación de carbono. Uso de la ATP y NADPH producido por las reacciones dependientes de la luz, los compuestos resultantes se reducen a triosa fosfato. De cada seis moléculas de fosfato de triosa producidos, uno se retira para formar más carbohidratos y cinco son "reciclados" de nuevo en el ciclo de regenerar el aceptante original de dióxido de carbono, RuBP.
Los primeros organismos fotosintéticos probablemente evolucionaron a principios de la la historia evolutiva de la vida. Los primeros organismos fotosintéticos probablemente utilizados agentes reductores tales como hidrógeno o sulfuro de hidrógeno como fuentes de electrones, en lugar de agua. Las cianobacterias que oxida el oxígeno en el agua apareció más tarde, y el exceso de oxígeno que producían condujo a la catástrofe de oxígeno, lo que finalmente hizo que la la evolución de la vida compleja posible. Hoy en día, la tasa promedio de captación de energía por fotosíntesis a nivel mundial es de aproximadamente 130 teravatios, que es cerca de seis veces más grande que el actual el consumo de energía de la civilización humana. En el proceso de los organismos fotosintéticos también convierten alrededor de 100 a 115.000 millones de toneladas métricas (es decir, 100-115 petagramos) de carbono en biomasa por año.
Visión de conjunto
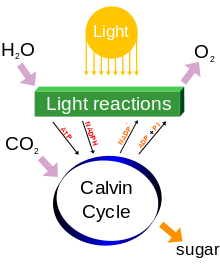
Los organismos fotosintéticos son Fotoautótrofos, lo que significa que son capaces de sintetizar los alimentos directamente a partir de dióxido de carbono y agua utilizando la energía de la luz. Sin embargo, no todos los organismos que utilizan la luz como fuente de energía llevar a cabo la fotosíntesis, ya fotoheterótrofos utilizan compuestos orgánicos, en lugar de dióxido de carbono, como fuente de carbono. En las plantas, algas y cianobacterias, la fotosíntesis libera oxígeno. Esto se llama la fotosíntesis oxigénica. Aunque hay algunas diferencias entre la fotosíntesis oxigénica en plantas , algas , y cianobacterias, el proceso en general es bastante similar en estos organismos. Sin embargo, hay algunos tipos de bacterias que llevan a cabo fotosíntesis anoxigénica, que consume dióxido de carbono, pero no libera oxígeno.
El dióxido de carbono se convierte en azúcares en un proceso llamado fijación de carbono. Fijación de carbono es un endotérmica reacción redox, por lo que la fotosíntesis necesita para suministrar tanto una fuente de energía para impulsar este proceso, y los electrones necesaria para convertir dióxido de carbono en un hidrato de carbono . Esta adición de los electrones es una reacción de reducción. En líneas generales y, en efecto, la fotosíntesis es el opuesto de la respiración celular, en el que la glucosa y otros compuestos se oxidan para producir dióxido de carbono y agua, y para liberar energía química exotérmica para conducir el organismo de metabolismo. Sin embargo, los dos procesos tienen lugar a través de una secuencia diferente de las reacciones químicas y en diferentes compartimentos celulares.
El general Por lo tanto, la ecuación para la fotosíntesis es:
- 2n CO 2 + 2n + DH 2 fotones → 2 (CH 2 O) n + 2n HACER
El dióxido de carbono + donador de electrones + luz de energía → carbohidratos + donador de electrones se oxida
En el agua la fotosíntesis oxigénica es el donador de electrones y, desde su hidrólisis libera oxígeno, la ecuación para este proceso es:
- 2n CO 2 + 4n H2O + fotones → 2 (CH 2 O) n + 2n O 2 + 2n H 2 O
- dióxido de carbono + agua + energía de la luz → carbohidratos + oxígeno + agua
A menudo, las moléculas de agua 2n se cancelan en ambos lados, obteniéndose:
- 2n CO 2 + 2n H2O + fotones → 2 (CH 2 O) n + 2n O 2
- dióxido de carbono + agua + energía de la luz → carbohidratos + oxígeno
Otros procesos de otros compuestos (tales como sustitutos arsenito) para el agua en el papel de electrones de suministro; por ejemplo, algunos microbios utilizan la luz solar para oxidar arsenito a arseniato: La ecuación para esta reacción es:
- CO 2 + (ASO 3 de 3) + fotones → (ASO 4 3-) + CO
- dióxido de carbono + arsenito + luz de energía → arseniato + monóxido de carbono (usado para construir otros compuestos en reacciones posteriores)
La fotosíntesis se produce en dos etapas. En la primera etapa, las reacciones dependientes de la luz o las reacciones de luz capturan la energía de la luz y la utilizan para hacer las moléculas de almacenamiento de energía ATP y NADPH. Durante la segunda etapa, las reacciones independientes de la luz usan estos productos para capturar y reducir el dióxido de carbono.
La mayoría de los organismos que utilizan la fotosíntesis para producir el uso de oxígeno la luz visible para hacerlo, aunque por lo menos tres utilización de onda corta infrarrojos o, más específicamente, la radiación rojo lejano.
Membranas fotosintéticas y orgánulos
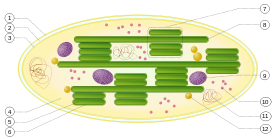
1. membrana externa
2. espacio intermembrana
3. membrana interna (1 + 2 + 3: sobre)
4. estroma (fluido acuoso)
5. lumen tilacoides (en el interior del tilacoide)
6. membrana tilacoide
7. granum (pila de tilacoides)
8. tilacoide (láminas)
9. almidón
10. ribosoma
11. ADN plastidial
12. plastoglobule (gota de lípidos)
En las bacterias fotosintéticas, las proteínas que recogen la luz para la fotosíntesis se incrustan dentro las membranas celulares, que es la configuración más sencilla estas proteínas están dispuestos. Sin embargo, esta membrana puede ser bien doblado en láminas cilíndricas llamadas tilacoides o amontonada en ronda vesículas llamadas membranas intracitoplasmáticos. Estas estructuras pueden llenar la mayor parte del interior de una célula, dando la membrana un área de superficie muy grande y por lo tanto aumentar la cantidad de luz que las bacterias pueden absorber.
En las plantas y algas, la fotosíntesis tiene lugar en orgánulos llamados cloroplastos. Una típica célula de la planta contiene alrededor de 10 a 100 cloroplastos. El cloroplasto está encerrado por una membrana. Esta membrana se compone de una membrana de fosfolípidos interior, una membrana externa de fosfolípidos, y un espacio intermembrana entre ellos. Dentro de la membrana es un fluido acuoso llamado el estroma. El estroma contiene pilas (Grana) de tilacoides, que son el sitio de fotosíntesis. Los tilacoides son discos aplanados, delimitadas por una membrana con un lumen o espacio tilacoide dentro de ella. El sitio de la fotosíntesis es la membrana tilacoide, que contiene integral y complejos de proteínas de membrana periféricas, incluyendo los pigmentos que absorben la energía de la luz, que forman los fotosistemas.
Las plantas absorben la luz principalmente mediante el pigmento clorofila, que es la razón de que la mayoría de las plantas tienen un color verde. Además de la clorofila, las plantas también utilizan pigmentos como carotenos y xantofilas. Las algas también utilizan la clorofila, pero varios otros pigmentos están presentes como ficocianina, carotenos, y xantofilas en algas verdes, ficoeritrina en algas rojas (Rhodophyta) y fucoxantina en algas marrones y diatomeas resultante en una amplia variedad de colores.
Estos pigmentos están incrustados en las plantas y algas en antena proteínas especiales. En tales proteínas todos los pigmentos están ordenados a trabajar bien juntos. Tal una proteína también se denomina complejo captador de luz.
Aunque todas las células en las partes verdes de una planta han cloroplastos, la mayor parte de la energía es capturada en el hojas, excepto en ciertas especies adaptadas a las condiciones de luz solar intensa y aridez, como muchos Euphorbia y Cactus especies, cuyo principal fotosintética órganos son sus tallos. Las células en los tejidos interiores de una hoja, llamada mesófilo, puede contener entre 450.000 y 800.000 cloroplastos por cada milímetro cuadrado de la hoja. La superficie de la hoja se recubre uniformemente con un resistente al agua- de cera cutícula que protege la hoja de excesiva evaporación del agua y disminuye la absorción de ultravioleta o azul luz para reducir el calentamiento . El transparente capa de la epidermis permite que la luz pase a través de la células del mesófilo en empalizada donde la mayoría de la fotosíntesis tiene lugar.
Reacciones luminosas
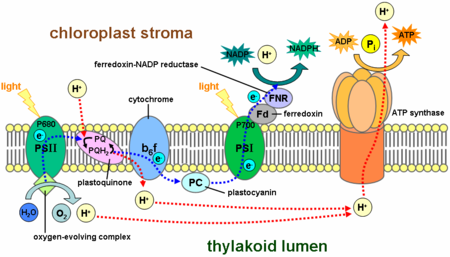
En el reacciones de luz, una molécula del pigmento clorofila absorbe un fotón y pierde un electrón . Este electrón se pasa a una forma modificada de la clorofila llamado feofitina, que pasa el electrón a una molécula de quinona, lo que permite el inicio de un flujo de electrones hacia abajo una cadena de transporte de electrones que conduce a una reducción de las NADP a NADPH. Además, esto crea una gradiente de protones a través de la membrana del cloroplasto; su disipación es utilizado por ATP sintasa para la síntesis concomitante de ATP . La molécula de clorofila recupera el electrón perdido a partir de un agua molécula a través de un proceso llamado fotólisis, que libera una dioxígeno (O 2) molécula. La ecuación general para las reacciones dependientes de la luz en las condiciones de flujo de electrones no cíclico en las plantas verdes es:
- 2 H 2 O 2 + NADP + + 3 ADP + 3 P i + iluminación → 2 NADPH + H + 2 + 3 + ATP O 2
No todo longitudes de onda de la luz pueden apoyar la fotosíntesis. El espectro de acción fotosintética depende del tipo de pigmentos accesorios presentes. Por ejemplo, en las plantas verdes, la espectro de acción se asemeja a la espectro de absorción para clorofilas y carotenoides con picos de violeta-azul y luz roja. En las algas rojas, el espectro de acción se superpone con el espectro de absorción de ficobilinas para la luz azul-verde rojo, lo que permite a estas algas que crecen en aguas más profundas que filtran las longitudes de onda más largas utilizadas por las plantas verdes. La parte no absorbida de la espectro de la luz es lo que le da su color fotosintética Microorganismos (por ejemplo, las plantas verdes, algas rojas, bacterias púrpuras) y es el menos eficaz para la fotosíntesis en los respectivos organismos.
Esquema Z
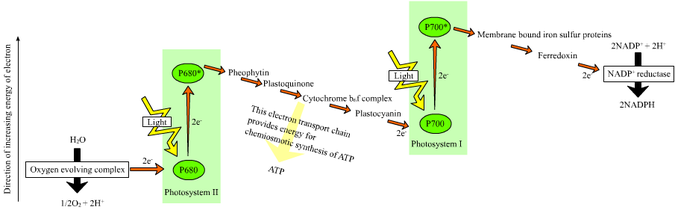
En las plantas, reacciones dependientes se producen en el membranas tilacoides de la cloroplastos y el uso de energía de luz para sintetizar ATP y NADPH. La reacción dependiente de la luz tiene dos formas: cíclicos y no cíclicos. En la reacción no cíclico, los fotones son capturados en la recolección de luz complejos de antena de fotosistema II por clorofila y otros pigmentos accesorios (véase el diagrama de la derecha). Cuando una molécula de clorofila en el centro de el centro de reacción del fotosistema II obtiene suficiente energía de excitación de los pigmentos de antena adyacentes, un electrón se transfiere a la molécula de aceptor de electrones primario, feofitina, a través de un proceso llamado separación de carga fotoinducida. Estos electrones son transportados a través de una cadena de transporte de electrones, el llamado Z-esquema que se muestra en el diagrama, que inicialmente funciones para generar una potencial quimiosmótica través de la membrana. Una ATP sintasa de la enzima utiliza el potencial quimiosmótica para producir ATP durante la fotofosforilación, mientras NADPH es un producto de la terminal de reacción redox en la Z-esquema. El electrón entra en una molécula de clorofila en Fotosistema I. El electrón es excitado por la luz absorbida por la fotosistema. Un segundo transportador de electrones acepta el electrón, que a su vez se transmite de bajar energías de receptores de electrones. La energía creada por los aceptores de electrones se utiliza para mover los iones de hidrógeno a través de la membrana tilacoide en el lumen. El electrón se utiliza para reducir la co-enzima NADP, que tiene funciones en la reacción de luz independiente. La reacción cíclica es similar a la de la no cíclico, pero difiere en la forma que sólo genera ATP, y no reducida de NADP (NADPH) se crea. La reacción cíclica se lleva a cabo sólo en el fotosistema I. Una vez que el electrón se desplaza del fotosistema, el electrón se transmite de las moléculas aceptoras de electrones y devuelve al fotosistema I, desde donde se emite, por lo tanto, la reacción cíclica nombre.
Fotolisis del agua
El NADPH es la principal agente reductor en los cloroplastos, proporcionando una fuente de electrones energéticos a otras reacciones. Su producción deja de clorofila con un déficit de electrones (oxidado), que se debe obtener de algún otro agente reductor. Los electrones excitados perdidos de clorofila en fotosistema I se sustituyen por la cadena de transporte de electrones por plastocianina. Sin embargo, desde fotosistema II incluye los primeros pasos de la Z-esquema, se requiere una fuente externa de electrones para reducir sus clorofila a moléculas oxidadas. La fuente de los electrones en el verde de las plantas y la fotosíntesis cianobacterias es el agua. Dos moléculas de agua se oxidan por cuatro reacciones de carga de separación sucesivas por el fotosistema II para proporcionar una molécula diatómica de oxígeno y cuatro de hidrógeno iones; el electrón cedido en cada paso se transfiere a un redox-activo residuo de tirosina, que luego reduce la clorofila-emparejado photoxidized una especie llamada P680 que sirve como el principal donante (impulsada por la luz) de electrones en el centro de reacción del fotosistema II. La oxidación del agua es catalizada en el fotosistema II por una estructura redox activo que contiene cuatro de manganeso iones y un ion de calcio; este compleja evolución de oxígeno se une a dos moléculas de agua y almacena los cuatro equivalentes oxidantes que se requieren para conducir la reacción de oxidación-agua. Fotosistema II es el único conocido biológica enzima que lleva a cabo esta oxidación del agua. Los iones de hidrógeno contribuyen a la potencial transmembrana quimiosmótica que conduce a la síntesis de ATP. El oxígeno es un producto de desecho de las reacciones dependientes de la luz, pero la mayoría de los organismos de la Tierra utilizan oxígeno para la respiración celular, incluyendo organismos fotosintéticos.
Fase oscura
Ciclo de Calvin
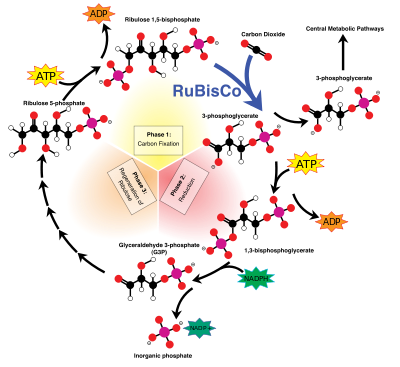
En el reacciones independientes de la luz (o "oscuros"), la enzima RuBisCO captura de CO 2 de la atmósfera y en un proceso que requiere la NADPH recién formado, llamado el ciclo de Calvin-Benson, libera azúcares de tres carbonos, que luego se combinan para formar la sacarosa y almidón. La ecuación general para las reacciones independientes de la luz en las plantas verdes es:
- 3 CO 2 + 9 + 6 ATP NADPH + 6 H + → C 3 H 6 O 3-fosfato + ADP + 9 8 P i + 6 + NADP + 3 H 2 O
Para ser más específicos, la fijación de carbono produce un producto intermedio, que se convierte a continuación en los productos finales de hidratos de carbono. Los esqueletos de carbono producido por la fotosíntesis se utilizan de diversas maneras para formar otros compuestos orgánicos, tales como el material de construcción celulosa, como precursores para los lípidos y aminoácidos biosíntesis, o como combustible en respiración celular. Este último se produce no sólo en las plantas, sino también en los animales cuando la energía de las plantas se pasa a través de una cadena de comida.
La fijación o la reducción de dióxido de carbono es un proceso en el que el dióxido de carbono se combina con un azúcar de cinco carbonos, ribulosa 1,5-bisfosfato (RuBP), para producir dos moléculas de un compuesto de tres carbonos, glicerato 3-fosfato (GP), también conocido como 3-fosfoglicerato (PGA). GP, en presencia de ATP y NADPH desde las etapas dependientes de la luz, se reduce a gliceraldehído 3-fosfato (G3P). Este producto también se conoce como 3-fosfogliceraldehido ( PGAL) o incluso como fosfato triosa. Triosa es un azúcar de 3 carbonos (ver hidratos de carbono ). La mayoría (5 de 6 moléculas) de la G3P producido se utiliza para regenerar RuBP por lo que el proceso puede continuar (ver Calvin-Benson ciclo). Los 1 de cada 6 moléculas de los fosfatos de triosa no "reciclados" a menudo se condensan para formar fosfatos de hexosa, que en última instancia producen sacarosa , almidón y celulosa. Los azúcares producidos durante carbono esqueletos de carbono rendimiento del metabolismo que pueden ser utilizados para otras reacciones metabólicas como la producción de aminoácidos y lípidos .
Mecanismos de carbono para concentrarse
En tierra
En condiciones calurosas y secas, las plantas cierran sus estomas para prevenir la pérdida de agua. En estas condiciones, CO 2 disminuirá, y el gas de oxígeno, producido por las reacciones luminosas de la fotosíntesis, disminuirá en el tallo, las hojas no, provocando un aumento de por la fotorrespiración actividad oxigenasa de ribulosa-1,5-bifosfato carboxilasa / oxigenasa y la disminución de la fijación de carbono. Algunas plantas han evolucionado mecanismos para aumentar la concentración de CO 2 en las hojas en estas condiciones.
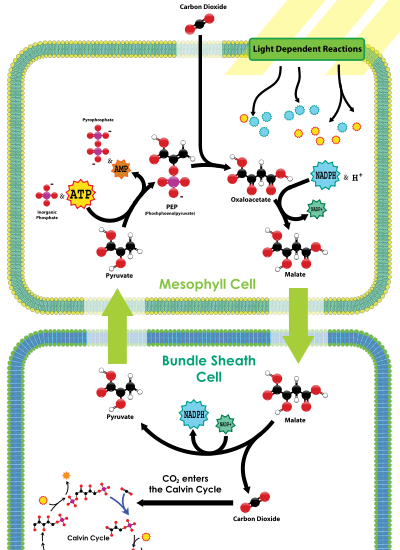
Plantas C 4 fijar químicamente dióxido de carbono en las células del mesófilo añadiéndolo a la molécula de tres carbonos fosfoenolpiruvato (PEP), una reacción catalizada por una enzima llamada PEP carboxilasa, creando el ácido orgánico de cuatro carbonos ácido oxaloacética. Ácido oxaloacético o malato sintetizado por este proceso se transloca luego a especializada células de la vaina del haz, donde la enzima Enzimas del ciclo de Rubisco y otra Calvino se encuentran, y donde CO2 liberados por descarboxilación de los ácidos de cuatro carbonos se fija entonces por la actividad RuBisCO al azúcar de tres carbonos Ácidos 3-fosfoglicérico. La separación física de la RuBisCO de las reacciones luminosas de generación de oxígeno reduce la fotorrespiración y aumenta la fijación de CO 2 y, por tanto, capacidad fotosintética de la hoja. C 4 plantas pueden producir más azúcar que C 3 plantas en condiciones de alta luz y la temperatura. Muchas plantas de cultivo importantes son C 4 plantas, incluyendo maíz, sorgo, caña de azúcar, y mijo. Las plantas que no utilizan PEP-carboxilasa en la fijación de carbono se denominan C 3 plantas debido a la reacción de carboxilación primaria, catalizada por la RuBisCO, produce el azúcar de tres carbonos ácidos 3-fosfoglicérico directamente en el ciclo de Calvin-Benson. Más del 90% de las plantas utilizan C3 fijación de carbono, frente al 3% que utilizan C 4 fijación de carbono.
Xerófitas, como los cactus y más suculentas, también utilizan PEP carboxilasa para capturar el dióxido de carbono en un proceso llamado Metabolismo ácido de las crasuláceas (CAM). En contraste con C 4 metabolismo, que separa físicamente la fijación de CO 2 a PEP al ciclo de Calvin, CAM separa temporalmente estos dos procesos. Plantas CAM tienen una anatomía hoja diferente de plantas C 3, y fijan el CO 2 en la noche, cuando sus estomas están abiertos. Plantas CAM almacenar el CO 2 en su mayoría en forma de ácido málico a través de carboxilación de fosfoenolpiruvato en oxaloacetato, que después se reduce a malato. La descarboxilación de malato durante el día libera CO 2 dentro de las hojas, lo que permite la fijación de carbono a 3-fosfoglicerato por RuBisCO. Dieciséis mil especies de plantas utilizan CAM.
En el agua
Las cianobacterias poseen carboxysomes, que aumentan la concentración de CO 2 alrededor de la RuBisCO para aumentar la tasa de fotosíntesis. Una enzima, anhidrasa carbónica, que se encuentra dentro de la Carboxisoma libera CO2 de los iones disueltos hidrocarbonato (HCO 3 -). Antes de que el CO 2 se difunde fuera se esponja rápidamente por RuBisCO, que se concentra en los carboxysomes. HCO 3 - iones están hechos de CO 2 fuera de la célula por otro de la anhidrasa carbónica y se bombean de forma activa en la célula por una proteína de membrana. Ellos no pueden atravesar la membrana a medida que se cargan, y dentro del citosol que girar de nuevo en CO 2 muy lentamente sin la ayuda de la anhidrasa carbónica. Esto hace que el HCO 3 - iones a acumularse dentro de la célula desde donde se difunden en los carboxysomes. Pyrenoids en algas y hornworts también actúan para concentrarse CO 2 alrededor rubisco.
Orden y cinética
El proceso global de la fotosíntesis se lleva a cabo en cuatro etapas:
| Etapa | Descripción | Escala de tiempo |
|---|---|---|
| 1 | La transferencia de energía en clorofila antena (membranas tilacoides) | femtosegundo a picosegundos |
| 2 | Transferencia de electrones en las reacciones fotoquímicas (membranas tilacoides) | picosegundos a nanosegundo |
| 3 | Cadena de transporte de electrones y la síntesis de ATP (membranas tilacoides) | microsegundo a milisegundo |
| 4 | La fijación de carbono y la exportación de productos estables | milisegundos para segundo |
Eficiencia

Plantas generalmente convierten la luz en energía química con una la eficiencia fotosintética de 3.6%. Eficiencia fotosintética plantas reales 'varía con la frecuencia de la luz que está siendo convertido, intensidad de la luz, la temperatura y la proporción de dióxido de carbono en la atmósfera, y puede variar de 0,1% a 8%. En comparación, paneles solares convierten la luz en energía eléctrica con una eficiencia de aproximadamente 6-20% para los paneles producidos en masa, y por encima de 40% en los dispositivos de laboratorio.
Sistemas de medición de la fotosíntesis no están diseñados para medir directamente la cantidad de luz absorbida por la hoja. Sin embargo, las curvas de respuesta de luz que sistemas como el producto LCpro-SD, no permiten comparaciones de la eficiencia fotosintética entre las plantas.
Evolución

Los primeros sistemas fotosintéticos, tales como los de verde y azufre púrpura y verde y bacterias del azufre púrpura, se cree que han sido anoxigénica, utilizando diversas moléculas como donadores de electrones. Chlorobi y morado se cree que han utilizado el hidrógeno y azufre como donador de electrones. Bacterias verdes del azufre utilizado diversos amino y otra ácidos orgánicos. Bacterias púrpuras del azufre utilizaron una variedad de moléculas orgánicas no específicas. El uso de estas moléculas es consistente con la evidencia geológica que la atmósfera era altamente reducida en ese momento .
Los fósiles de lo que se cree que son organismos fotosintéticos filamentosos han sido datados en 3,4 millones de años.
La principal fuente de oxígeno en la atmósfera es fotosíntesis oxigénica, y su primera aparición se refiere a veces como la catástrofe oxígeno. La evidencia geológica sugiere que la fotosíntesis oxigénica, como el que en cianobacterias, se hizo importante durante el Era Paleoproterozoic Hace alrededor de 2 millones de años. Fotosíntesis Moderno de plantas y procariotas fotosintéticos es más oxigénica. Fotosíntesis oxigénica utiliza el agua como un donador de electrones, que es oxidada al oxígeno molecular (O 2) en el centro de reacción fotosintética .
Simbiosis y el origen de los cloroplastos
Varios grupos de animales han formado relaciones simbióticas con algas fotosintéticas. Estos son más comunes en corales, esponjas y anémonas de mar. Se presume que esto es debido a la particularmente simple planes corporales y grandes áreas de superficie de estos animales en comparación con sus volúmenes. Además, unos pocos marina moluscos Elysia viridis y Elysia chlorotica también mantienen una relación simbiótica con los cloroplastos capturan de las algas en su dieta y luego almacenan en sus cuerpos. Esto permite que los moluscos para sobrevivir únicamente por la fotosíntesis durante varios meses a la vez. Algunos de los genes de la planta núcleo de la célula se han de transferirse a las babosas, de modo que los cloroplastos se pueden suministrar con proteínas que necesitan para sobrevivir.
Una forma aún más cerca de la simbiosis puede explicar el origen de los cloroplastos. Los cloroplastos tienen muchas similitudes con bacterias fotosintéticas, incluyendo una circular cromosoma, de tipo procariota ribosomas y proteínas similares en el centro de reacción fotosintética. La teoría endosimbiótica sugiere que las bacterias fotosintéticas fueron adquiridas (por endocitosis) por los primeros eucariotas células para formar las primeras plantas células. Por lo tanto, los cloroplastos pueden ser bacterias fotosintéticas que se adaptaron a la vida en el interior de las células vegetales. Al igual que las mitocondrias , los cloroplastos todavía poseen su propio ADN, separada de la ADN nuclear de sus células huésped de la planta y los genes de este ADN del cloroplasto se asemejan a los de cianobacterias. ADN en los códigos de cloroplastos para proteínas redox tales como centros de reacción fotosintéticos. La Corr hipótesis propone que se requiere esta Co -ubicación de I edox R eglamento.
Las cianobacterias y la evolución de la fotosíntesis
La capacidad bioquímica de utilizar el agua como la fuente de electrones en la fotosíntesis evolucionó una vez, en una ancestro común de los existentes cianobacterias. El registro geológico indica que este acontecimiento transformador tuvo lugar temprano en la historia de la Tierra, por lo menos hace 2450-2320 millones de años (Ma), y, se especula, mucho antes. Las pruebas disponibles de los estudios de geobiológicos Arcaico (> 2.500 Ma) rocas sedimentarias indica que la vida existió 3.500 Ma, pero la cuestión de cuándo la fotosíntesis oxigénica evolucionó todavía sin respuesta. Una ventana paleontológico clara sobre cianobacterias evolución abrió unos 2000 Ma, revelando una biota diversa de ya-azul-verdes. Las cianobacterias se mantuvo director los productores primarios de todo el Proterozoico (2500-543 Ma), en parte porque la estructura redox de los océanos favoreció fotoautótrofos capaces de la fijación del nitrógeno . Las algas verdes se unió azul-verdes como los principales productores primarios en plataformas continentales cerca del final de la Proterozoico, pero sólo con el Mesozoico (251-65 Ma) radiaciones de dinoflagelados, cocolitofóridos, y diatomeas hicieron la producción primaria en aguas de la plataforma marina tomar forma moderna. Las cianobacterias siendo fundamental para los ecosistemas marinos como productores primarios en giros oceánicos, como agentes de la fijación biológica de nitrógeno y, en forma modificada, como el plástidos de algas marinas.
Un estudio de 2010 realizado por investigadores de La Universidad de Tel Aviv descubrió que el Hornet Oriental (Vespa orientalis) convierte la luz solar en energía eléctrica mediante un pigmento llamado xantopterina. Esta es la primera evidencia científica de un miembro del reino animal participar en la fotosíntesis.
Descubrimiento
Aunque algunos de los pasos en la fotosíntesis no están todavía completamente entendido, la ecuación general fotosintética se conoce desde el siglo 19.
Jan van Helmont comenzó la investigación del proceso en la mitad del siglo 17 cuando midió cuidadosamente la masa de la tierra usada por una planta y la masa de la planta a medida que crecía. Después de notar que la masa de suelo cambió muy poco, se planteó la hipótesis de que la masa de la planta en crecimiento debe venir del agua, la única sustancia añadió a la planta en maceta. Su hipótesis fue parcialmente exacta - gran parte de la masa de ganado también viene de dióxido de carbono, así como el agua. Sin embargo, este fue un punto de señalización a la idea de que la mayor parte de una planta es biomasa proviene de las entradas de la fotosíntesis, no el propio suelo.
Joseph Priestley, un químico y ministro, descubrió que, cuando se aisló un volumen de aire bajo un frasco invertido, y se quema una vela en ella, la vela se quema muy rápidamente, mucho antes de que se quedó sin cera. Descubrió, además, que un ratón podría igualmente "lesionar" aire. Luego mostró que el aire que había sido "herido" por la vela y el ratón podría ser restaurado por una planta.
En 1778, Jan Ingenhousz, médico de la corte a la austríaca emperatriz, repitió los experimentos de Priestley. Se descubrió que era la influencia de la luz solar sobre la planta que podrían causar que se revive un ratón en una cuestión de horas.
En 1796, Jean Senebier, un suizo pastor, botánico y naturalista, demostró que las plantas verdes consumen dióxido de carbono y liberan oxígeno bajo la influencia de la luz. Poco después, Nicolas-Théodore de Saussure mostró que el aumento de la masa de la planta a medida que crece no sólo podría ser debido a la absorción de CO 2, sino también a la incorporación de agua. Por lo tanto, se describe la reacción básica por la que la fotosíntesis se utiliza para producir alimentos (tales como glucosa).
Cornelis Van Niel hizo descubrimientos clave que explican la química de la fotosíntesis. Estudiando bacterias del azufre púrpura y bacterias verdes él era el primer científico en demostrar que la fotosíntesis es un dependiente de la luz reacción redox, en la que el hidrógeno reduce el dióxido de carbono.
Robert Emerson descubrió dos reacciones luminosas de la productividad pruebas de planta utilizando diferentes longitudes de onda de la luz. Con el rojo solo, se suprimieron las reacciones luminosas. Cuando azul y rojo fueron combinadas, la salida fue mucho más sustancial. Por lo tanto, había dos fotosistemas, una absorción de hasta 600 nm longitud de onda, la otra hasta 700 nm. El primero es conocido como PSII, este último es PSI. PSI sólo contiene clorofila a, PSII contiene clorofila principalmente una con la mayor parte de la clorofila b disponibles, entre otro pigmento. Estos incluyen ficobilinas, que son los pigmentos rojos y azules de algas de color rojo y azul respectivamente, y fucoxanthol para las algas pardas y diatomeas. El proceso es más productivo cuando la absorción de cuantos son iguales tanto en el PSII y PSI, asegurando que la energía de entrada del complejo de la antena se divide entre el PSI y PSII sistema, que a su vez impulsa la fotoquímica.

Robert Hill pensó que un complejo de reacciones que consisten de un compuesto intermedio de citocromo b 6 (ahora un plastoquinona), otra es de citocromo f para un paso en los mecanismos de generación de hidratos de carbono. Estos están unidos por plastoquinona, que requiere energía para reducir el citocromo f porque es un reductor suficiente. Otros experimentos para demostrar que el oxígeno se desarrolló durante la fotosíntesis de las plantas verdes vino del agua, se llevaron a cabo por Hill en 1937 y 1939. Demostró que aislado cloroplastos emiten oxígeno en presencia de agentes reductores no naturales como hierro oxalato, ferricianuro o benzoquinona después de la exposición a la luz. La reacción de Hill es la siguiente:
- 2 H 2 O + 2 A + (luz, cloroplastos) → 2 AH 2 + O 2
donde A es el aceptor de electrones. Por lo tanto, a la luz, el aceptor de electrones se reduce y se desprende oxígeno.
Samuel Ruben y Martin Kamen utilizado isótopos radiactivos para determinar que el oxígeno liberado en la fotosíntesis vino del agua.
Melvin Calvin y Andrew Benson, junto con James Bassham, dilucidó el camino de la asimilación de carbono (el ciclo de reducción de carbono fotosintético) en las plantas. El ciclo de reducción de carbono se conoce como el Ciclo de Calvin, que ignora la contribución de Bassham y Benson. Muchos científicos se refieren al ciclo que el ciclo de Calvin-Benson, Benson-Calvin, y algunos incluso lo llaman el ciclo de Calvin-Benson-Bassham (o CBB).
Científico ganador del Premio Nobel Rudolph A. Marcus fue capaz de descubrir la función y la importancia de la cadena de transporte de electrones.
Otto Heinrich Warburg y Dean Burk descubrió la reacción de la fotosíntesis I-cuántico que divide el CO 2, activado por la respiración.
Louis NM Duysens yEne Amesz descubrieron que la clorofila a absorbe una luz, oxidar citocromo f, clorofila a (y otros pigmentos) absorberá otra luz, pero reducirá este mismo citocromo oxidada, indicando las dos reacciones de luz son en serie.
Factores

Hay tres factores principales que afectan a la fotosíntesis y varios factores corolario. Los tres principales son:
- Luz irradiancia y longitud de onda
- Dióxido de carbono concentración
- Temperatura.
Intensidad de la luz (irradiación), longitud de onda y de la temperatura
En el siglo 20, Frederick Blackman yGabrielle Matthaei investigaron los efectos de la intensidad de luz (irradiancia) y la temperatura sobre la velocidad de asimilación de carbono.
- A temperatura constante, la tasa de asimilación de carbono varía con la irradiancia, inicialmente aumenta a medida que aumenta la irradiancia. Sin embargo, en mayor irradiancia, esta relación ya no se sostiene y la tasa de asimilación de carbono alcanza una meseta.
- En irradiación constante, la tasa de asimilación de carbono aumenta a medida que se aumenta la temperatura en un intervalo limitado. Este efecto se observó sólo en los altos niveles de irradiación. A baja irradiancia, el aumento de la temperatura tiene poca influencia en la tasa de asimilación de carbono.
Estos dos experimentos ilustran puntos vitales: En primer lugar, a partir de la investigación se sabe que, en general, las reacciones fotoquímicas no se ven afectados por la temperatura . Sin embargo, estos experimentos muestran claramente que la temperatura afecta a la tasa de asimilación de carbono, así que debe haber dos conjuntos de reacciones en el proceso completo de asimilación de carbono. Estos son, por supuesto, la fase dependiente de la luz 'fotoquímico "y la etapa de luz independiente, dependiente de la temperatura. En segundo lugar, los experimentos de Blackman ilustran el concepto de factores limitantes. Otro factor limitante es la longitud de onda de la luz. Las cianobacterias, que residen varios metros bajo el agua, no puede recibir las longitudes de onda correctas requeridas para causar separación de carga fotoinducida en pigmentos fotosintéticos convencionales. Para combatir este problema, una serie de proteínas con diferentes pigmentos rodean el centro de reacción. Esta unidad se llama un ficobilisoma.
Niveles de dióxido de carbono y la fotorrespiración
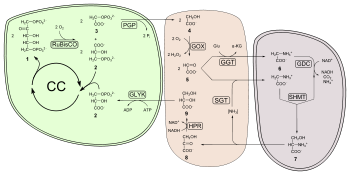
Como las concentraciones de dióxido de carbono aumentan, la velocidad a la que los azúcares son hechas por las reacciones independientes de la luz aumenta hasta limitado por otros factores. RuBisCO, la enzima que captura el dióxido de carbono en las reacciones independientes de la luz, tiene una afinidad de unión tanto para el dióxido de carbono y oxígeno. Cuando la concentración de dióxido de carbono es alto, RuBisCO se fijan el dióxido de carbono. Sin embargo, si la concentración de dióxido de carbono es baja, RuBisCO se unirá oxígeno en lugar de dióxido de carbono. Este proceso, llamado fotorrespiración, utiliza la energía, pero no produce azúcares.
Actividad oxigenasa RuBisCO es desventajoso para las plantas por varias razones:
- Uno de los productos de la actividad oxigenasa es phosphoglycolate (2 carbono) en lugar de 3-fosfoglicerato (3 de carbono). Phosphoglycolate no puede ser metabolizado por el ciclo de Calvin-Benson y representa pérdida de carbono del ciclo. Una alta actividad oxigenasa, por lo tanto, drena los azúcares que se requieren para reciclar ribulosa 5-bisfosfato y para la continuación de la ciclo de Calvin-Benson.
- Phosphoglycolate se metaboliza rápidamente a glicolato que es tóxico para una planta en una concentración alta; inhibe la fotosíntesis.
- Rescate glicolato es un proceso energéticamente costoso que utiliza la vía glicolato, y sólo 75% del carbono se devuelve al ciclo de Calvin-Benson como 3-fosfoglicerato. Las reacciones también producen amoníaco (NH 3 ), que es capaz de difundirse fuera de la planta, lo que lleva a una pérdida de nitrógeno.
- Un resumen muy simplificado es:
- 2 glicolato + ATP → 3-fosfoglicerato + dióxido de carbono + ADP + NH3
La vía de salvamento para los productos de la actividad oxigenasa RuBisCO se conoce más comúnmente comofotorrespiración, ya que se caracteriza por el consumo de oxígeno dependiente de la luz y la liberación de dióxido de carbono.