


Évolution
À propos de ce écoles sélection Wikipedia
Cette sélection écoles a été choisi par SOS Enfants pour les écoles dans le monde en développement ne ont pas accès à Internet. Il est disponible en téléchargement intranet. Avec enfants SOS vous pouvez choisir de parrainer des enfants dans plus de cent pays
L'évolution est la variation de la hérité caractéristiques du biologique populations plus successive générations. Les processus évolutifs donnent lieu à la diversité à tous niveau d'organisation biologique, y compris les espèces , les organismes individuels et des molécules telles que l'ADN et les protéines .
La vie sur Terre a évolué à partir d'un universelle ancêtre commun il ya environ 3,8 milliards d'années. Répété spéciation et la divergence de la vie peut être déduit de ensembles partagés de caractères biochimiques et morphologiques ou par Shared des séquences d'ADN. Ces les traits et des séquences homologues sont plus similaires entre espèces qui partagent un ancêtre commun plus récent, et peuvent être utilisées pour reconstruire histoires évolutives, en utilisant deux espèces existantes et le registre fossile . Les modèles existants de la biodiversité ont été façonnées à la fois par la spéciation et en extinction .
Charles Darwin et Alfred Wallace a été le premier à formuler un argument scientifique pour la théorie de l'évolution au moyen de la sélection naturelle . Évolution par sélection naturelle est un processus qui est déduit de trois faits sur les populations: 1) plus de descendants sont produits qu'il ne en peut survivre, 2) traits varient entre les individus, conduisant à différents taux de survie et la reproduction, et 3) les différences de traits sont héréditaires. Ainsi, lorsque les membres d'une population meurent, ils sont remplacés par le progéniture des parents qui étaient mieux adapté pour survivre et se reproduire dans le environnement dans lequel la sélection naturelle a eu lieu. Ce processus crée et préserve des traits qui sont apparemment équipée pour le rôles fonctionnels qu'ils effectuent. La sélection naturelle est la seule cause connue de adaptation, mais pas la seule cause connue de l'évolution. Autres, les causes de non adaptatifs évolution inclut mutation et dérive génétique.
Au début du 20e siècle, la génétique était intégré avec la théorie darwinienne de l'évolution par la sélection naturelle par la discipline des la génétique des populations. L'importance de la sélection naturelle comme une cause de l'évolution a été accepté dans d'autres branches de la biologie . Notions plus, détenues précédemment sur l'évolution, tels que orthogenèse et «Progrès» est devenu obsolète. Les scientifiques continuent d' étudier divers aspects de l'évolution en formant et tester des hypothèses, de la construction théories scientifiques, à l'aide données d'observation et performant expériences à la fois la terrain et en laboratoire. Biologistes conviennent que la descendance avec modification est l'un des plus établie de manière fiable faits de la science. Découvertes en biologie évolutive ont eu un impact significatif non seulement dans les branches traditionnelles de la biologie, mais aussi dans d'autres disciplines (par exemple, anthropologie et la psychologie) et sur la société dans son ensemble.
Histoire de la pensée évolutionniste
La proposition selon laquelle un type d'animal pouvait descendre d'un animal d'un autre type remonte à une partie du premier philosophes présocratiques grecs, tels que Anaximandre et Empédocle. Contrairement à ceux-ci vues matérialistes, Aristote compris toutes les choses naturelles, non seulement vivant des choses, comme étant imparfaite actualisations de différentes possibilités naturelles fixes, appelés " formes "," idées ", ou (dans les traductions latines)« espèce ». Cela faisait partie de son téléologique de compréhension la nature dans laquelle toutes les choses ont un rôle destiné à jouer dans un divin ordre cosmique. Le Romain poète et philosophe Titus Lucrèce a proposé la possibilité de changements évolutifs d'organismes. Variations de cette idée est devenu le standard de la compréhension du Moyen Age , et ont été intégrés dans l'apprentissage chrétien, mais Aristote ne exigeaient pas que les vrais types d'animaux correspondaient d'une pour une des formes métaphysiques exactes, et en particulier a donné des exemples de la façon dont de nouveaux types de êtres vivants pourrait venir d'être.
Au 17ème siècle, la nouvelle méthode de la science moderne a rejeté l'approche d'Aristote, et a demandé des explications des phénomènes naturels en termes de lois de la nature qui étaient les mêmes pour toutes les choses visibles, et ne ont pas besoin d'assumer des catégories naturelles fixes, ni aucun ordre cosmique divine. Mais cette nouvelle approche a été lent à prendre racine dans les sciences biologiques, qui est devenu le dernier bastion de la notion de types naturels fixes. John Ray a utilisé une des conditions préalablement plus générales pour les types naturels fixes, «espèce», à appliquer à des types animales et végétales, mais contrairement à Aristote il strictement identifié chaque type de chose vivante comme une espèce, et a proposé que chaque espèce peuvent être définis par les caractéristiques qui se perpétuent chaque génération. Ces espèces ont été conçus par Dieu, mais montrant les différences causées par les conditions locales. La classification biologique présenté par Carolus Linnaeus en 1735 ont aussi consulté espèces fixe selon un plan divin.

Autres naturalistes de cette époque ont spéculé sur le changement évolutif des espèces au fil du temps selon des lois naturelles. Maupertuis a écrit en 1751 des modifications naturelles survenant lors de la reproduction et de l'accumulation au fil des générations pour produire de nouvelles espèces. Buffon a suggéré que les espèces pourraient dégénérer en différents organismes, et Erasmus Darwin a proposé que tous les animaux à sang chaud pourraient descendent d'un seul micro-organisme (ou "filament"). Le premier schéma de l'évolution à part entière était "Transmutation" la théorie de 1809 de Lamarck qui prévoyait la génération spontanée produire continuellement les formulaires simples de la vie développé une plus grande complexité dans les lignées parallèles avec une tendance progressive inhérente, et que sur le plan local, ces lignées adapté à l'environnement par des changements héritant causés par l'utilisation ou la désuétude parents. (Ce dernier processus a été appelé plus tard lamarckisme.) Ces idées ont été condamnés par les naturalistes établies que la spéculation manque de soutien empirique. En particulier Georges Cuvier a insisté pour que les espèces ne étaient pas liés et fixe, leurs similitudes reflétant la conception divine pour les besoins fonctionnels. Dans l'intervalle, les idées de Ray de dessein bienveillant avaient été développé par William Paley dans un la théologie naturelle qui a proposé des adaptations complexes comme preuve de la conception divine, et a été admiré par Charles Darwin.
La rupture critique de la notion d'espèces fixes en biologie a commencé avec la théorie de l'évolution par la sélection naturelle, qui a été formulée par Charles Darwin. Partie influencé par Essai sur le principe de population par Thomas Robert Malthus , Darwin a noté que la croissance de la population conduirait à une «lutte pour l'existence" où les variations favorables pourraient prévaloir comme d'autres ont péri. Chaque génération, de nombreux descendants ne survivent pas à un âge de la reproduction en raison de ressources limitées. Cela pourrait expliquer la diversité des animaux et des plantes à partir d'un ancêtre commun à travers le travail des lois naturelles qui travaillent le même pour tous les types de chose. Darwin développait sa théorie de la « sélection naturelle »de 1838 et jusqu'au Alfred Russel Wallace lui a envoyé une théorie similaire en 1858. Les deux hommes ont présenté leur documents distincts à la Linnean Society of London. À la fin de 1859, la publication de Darwin sur l'origine des espèces explique la sélection naturelle en détail et d'une manière qui a conduit à une plus large acceptation de L'évolution darwinienne. Thomas Henry Huxley appliqué les idées de Darwin à l'homme, en utilisant la paléontologie et anatomie comparée de fournir des preuves solides que les humains et les grands singes partagent un ancêtre commun. Certains ont été perturbés par cette disposition parce qu'elle implique que les humains ne ont pas une place particulière dans l'univers.
Mécanismes précis de l'héritabilité de la reproduction et l'origine de nouveaux traits sont restés un mystère. À cette fin, Darwin a développé sa théorie provisoire de pangenèse. En 1865, Gregor Mendel a rapporté que les traits ont été hérités d'une manière prévisible par l'assortiment indépendant et la ségrégation des éléments (plus tard connu sous le nom gènes). Les lois de Mendel sur l'hérédité finalement supplantés plus de la théorie de pangenèse de Darwin. Août Weismann a fait la distinction importante entre cellules germinales (spermatozoïdes et ovules) et les cellules somatiques du corps, ce qui démontre que l'hérédité passe à travers la ligne germinale seulement. Hugo de Vries relié la théorie de Darwin à pangenèse de germes / cellule soma distinction de Weismann et a proposé que les pangènes de Darwin ont été concentrés dans le noyau cellulaire quand elle est exprimée et qu'ils puissent se déplacer dans la cytoplasme de modifier la structure des cellules. De Vries est également l'un des chercheurs qui ont fait des travaux de Mendel bien connu, croyant que mendélienne correspondent au transfert de variations héréditaires long de la lignée germinale. Pour expliquer comment les nouvelles variantes proviennent, De Vries a développé une la théorie de la mutation qui conduit à une rupture temporaire entre ceux qui ont accepté l'évolution darwinienne et biométriciens qui alliés avec de Vries. Au tournant du 20e siècle, des pionniers dans le domaine de la génétique des populations, tels que JBS Haldane, Sewall Wright, et Ronald Fisher, a jeté les bases de l'évolution sur une philosophie statistique solide. La fausse contradiction entre la théorie de Darwin, des mutations génétiques et l'hérédité mendélienne a donc réconcilié.
Dans les années 1920 et 1930 une la synthèse évolutionniste moderne relié sélection naturelle, la théorie de mutation, et l'hérédité mendélienne dans une théorie unifiée qui se applique généralement à toute branche de la biologie. La synthèse moderne a été en mesure d'expliquer les tendances observées à travers les espèces dans les populations, à travers transitions fossiles dans la paléontologie, et mécanismes cellulaires complexes, même dans la biologie du développement. La publication de la structure de l'ADN par James Watson et Francis Crick en 1953 a démontré une base physique pour héritage. La biologie moléculaire a amélioré notre compréhension de la relation entre le génotype et le phénotype. Les progrès ont également été faites dans phylogénétique la systématique, la cartographie de la transition de traits dans un cadre comparatif et vérifiable par la publication et l'utilisation de arbres évolutifs. En 1973, le biologiste évolutionniste Theodosius Dobzhansky a écrit que «rien en biologie n'a de sens qu'à la lumière de l'évolution", car il a mis en lumière les relations de ce qui semblait d'abord faits disjoints d'histoire naturelle en un tout cohérent corps explicatif de connaissances qui décrit et prédit de nombreux faits observables sur la vie sur cette planète.
Depuis lors, la synthèse moderne a été étendue pour expliquer les phénomènes biologiques dans la pleine échelle et intégrative de la hiérarchie biologique, des gènes aux espèces. Cette extension a été surnommé " éco-evo-devo ".
Hérédité
Evolution dans les organismes se produit par des changements dans héréditaires traits - des caractéristiques particulières d'un organisme. Chez l'homme, par exemple, la couleur des yeux est une caractéristique héritée et un individu peut hériter de la «trait brun yeux" d'un de leurs parents. Caractères héréditaires sont contrôlées par les gènes et l'ensemble complet de gènes dans un organisme de est appelé son génome génotype.
L'ensemble des caractéristiques observables qui composent la structure et le comportement d'un organisme est appelé le phénotype. Ces caractéristiques proviennent de l'interaction de son génotype avec le environnement. En conséquence, de nombreux aspects du phénotype d'un organisme ne sont pas héritées. Par exemple, la peau bronzée provient de l'interaction entre le génotype et le soleil d'une personne; Ainsi, bronzage ne sont pas transmis aux enfants des gens. Cependant, certaines personnes bronzent plus facilement que d'autres, en raison de différences dans leur génotype; un exemple frappant des gens avec le caractère héréditaire de l'albinisme , qui ne bronzent pas du tout et sont très sensibles à les coups de soleil.
Traits héréditaires sont transmis d'une génération à l'autre par l'intermédiaire de l'ADN , une molécule qui code l'information génétique. L'ADN est une longue polymère composé de quatre types de bases. La séquence de bases le long d'une molécule d'ADN particulière spécifier l'information génétique, d'une manière similaire à une séquence de lettres observe définissant une phrase. Avant qu'une cellule se divise, l'ADN est copié, de sorte que chacune des deux cellules résultantes hériteront la séquence d'ADN. Portions d'une molécule d'ADN qui spécifient une unité fonctionnelle unique sont appelés gènes; gènes différents ont des séquences de bases. Dans les cellules , les longues mèches de forme condensée structures d'ADN appelés chromosomes. La localisation spécifique d'une séquence d'ADN dans un chromosome est connu comme un locus. Si la séquence d'ADN à un locus varie selon les individus, les différentes formes de cette séquence sont appelés allèles. séquences d'ADN peuvent changer par le biais mutations, la production de nouveaux alleles. Si une mutation se produit dans un gène, le nouvel allèle peut affecter la caractéristique que les contrôles de gènes, la modification du phénotype de l'organisme. Toutefois, bien que cette correspondance simple entre un allèle d'un caractère et fonctionne dans certains cas, la plupart des caractères sont plus complexes et sont contrôlés par de multiples gènes qui interagissent.
Des découvertes récentes ont confirmé d'importants exemples de changements héréditaires qui ne peuvent pas être expliquées par des changements à la séquence de nucléotides dans l'ADN. Ces phénomènes sont considérés comme systèmes de transmission épigénétique. ADN marquage méthylation chromatine, boucles métaboliques autonome, silençage génique par l'interférence ARN et la tridimensionnel conformation des protéines (tels que les prions ) sont des domaines où les systèmes de transmission épigénétique ont été découverts au niveau organismique. Biologistes du développement suggèrent que les interactions complexes réseaux génétiques et la communication entre les cellules peuvent entraîner des variations héréditaires qui peuvent sous-tendent certains des mécanismes de plasticité du développement et la canalisation. Héritabilité peut également se produire à échelles encore plus grandes. Par exemple, l'héritage écologique dans le processus de la construction de niche est définie par les activités régulières et répétées des organismes dans leur environnement. Cela génère des effets qui modifient et se nourrissent de nouveau dans le régime des générations ultérieures de sélection un héritage. Descendants héritent des gènes ainsi que les caractéristiques environnementales générées par les actions écologiques des ancêtres. D'autres exemples de l'héritabilité de l'évolution qui ne sont pas sous le contrôle direct de gènes comprennent l'héritage de traits culturels et symbiogenèse.
Variation


De l'organisme individuel résultats phénotypiques de son à la fois génotype et l'influence de la environnement, il a vécu. Une partie substantielle de la variation des phénotypes dans une population est causé par les différences entre leurs génotypes. Le la synthèse évolutionniste moderne définit l'évolution comme le changement au fil du temps dans cette variation génétique. La fréquence d'un allèle particulier sera plus ou moins fréquente par rapport à d'autres formes de ce gène. Variation disparaît quand un nouvel allèle atteint le point de fixation - soit quand il disparaît de la population ou remplace l'allèle ancestrale entièrement.
La sélection naturelle ne fera que provoquer évolution se il ya assez la variation génétique dans une population. Avant la découverte de La génétique mendélienne, une hypothèse commune était héritage mélange. Mais avec le mélange l'héritage, la variance génétique serait rapidement perdue, rendant l'évolution par la sélection naturelle invraisemblable. Le Principe de Hardy-Weinberg fournit la solution à la façon dont la variation est maintenue dans une population Hérédité mendélienne. Les fréquences des alleles (variations dans un gène) resteront constants en l'absence de sélection, la mutation, la migration et la dérive génétique.
Variation provient de des mutations dans matériel génétique, remaniement de gènes par le biais la reproduction sexuée et la migration entre les populations ( flux de gènes). Malgré l'introduction constante de nouvelle variation par mutation et le flux génétique, la plupart des génome d'une espèce est identique chez tous les individus de la même espèce. Cependant, même de petites différences dans le génotype peuvent conduire à des différences spectaculaires dans phénotype: par exemple, les chimpanzés et les humains diffèrent que dans environ 5% de leurs génomes.
Mutation
Les mutations sont des changements dans la séquence d'ADN du génome d'une cellule. Lorsque mutations se produisent, ils peuvent soit avoir aucun effet, modifier la produit d'un gène, ou le gène empêcher de fonctionner. Basé sur des études chez la mouche Drosophila melanogaster , il a été suggéré que si une mutation modifie une protéine produite par un gène, ce sera probablement nuisible, avec environ 70% de ces mutations ayant des effets nuisibles, et le reste étant soit neutre ou faiblement bénéfique.
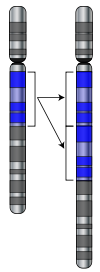
Les mutations peuvent impliquer de grandes sections de devenir un chromosome dupliqué (généralement par recombinaison génétique) qui peut introduire des copies supplémentaires d'un gène dans un génome. Des copies supplémentaires de gènes sont une source majeure de la matière première nécessaire pour les nouveaux gènes d'évoluer. Ceci est important car la plupart des nouveaux gènes évoluent à l'intérieur familles de gènes de gènes préexistants qui partagent des ancêtres communs. Par exemple, l'œil humain utilise quatre gènes de réaliser des structures sensibles à la lumière: trois pour la vision des couleurs et une pour la vision de nuit; tous les quatre sont les descendants d'un gène ancestral unique.
De nouveaux gènes peuvent être générés à partir d'un gène ancestral quand un Duplicata mute et acquiert une nouvelle fonction. Ce processus est plus facile une fois un gène a été dupliqué car elle augmente la la redondance du système; un gène de la paire peut acquérir une nouvelle fonction tandis que l'autre copie continue à remplir sa fonction initiale. D'autres types de mutations peuvent même générer entièrement nouveaux gènes non codante de l'ADN précédemment.
La génération de nouveaux gènes peut également impliquer les petites parties de plusieurs gènes étant dupliqués, avec ces fragments recombiner ensuite pour former de nouvelles combinaisons avec de nouvelles fonctions. Lorsque de nouveaux gènes sont assemblés à partir de pièces pré-brassage existantes, domaines agissent comme des modules indépendants avec des fonctions simples, qui peuvent être mélangés ensemble pour produire de nouvelles combinaisons avec des fonctions nouvelles et complexes. Par exemple, polykétides synthases sont grandes enzymes qui font antibiotiques; ils contiennent jusqu'à cent domaines indépendants qui catalysent chaque étape dans le processus global, comme une étape dans une chaîne de montage.
Sexe et la recombinaison
Dans les organismes asexués, les gènes sont hérités ensemble, ou liés, car ils ne peuvent pas se mélanger avec les gènes d'autres organismes lors de la reproduction. En revanche, la progéniture de organismes sexuelles contiennent des mélanges aléatoires des chromosomes de leurs parents qui sont produites par assortiment indépendant. Dans un processus similaire appelé recombinaison homologue, organismes sexuelles échanger ADN entre deux chromosomes correspondant. Recombinaison et réassortiment ne modifient pas les fréquences des allèles, mais qui changent allèles sont associés les uns aux autres, produire une descendance avec de nouvelles combinaisons d'allèles. Sex augmente généralement la variation génétique et peut augmenter le taux de l'évolution.
Le flux de gènes
Le flux de gènes est l'échange de gènes entre les populations et entre espèces. Il peut donc être une source de variation qui est nouveau pour une population ou à une espèce. Le flux de gènes peut être causée par la circulation des personnes entre les populations distinctes d'organismes, comme on pourrait être causé par le mouvement de la souris entre les terres et les populations côtières, ou le mouvement de pollen entre les tolérants aux métaux lourds et les populations sensibles de métaux lourds de graminées.
Le transfert de gènes entre les espèces comprend la formation de organismes hybrides et transfert horizontal de gènes. Le transfert de gène horizontal est le transfert de matériel génétique d'un organisme à un autre organisme qui ne est pas sa progéniture; ce est plus fréquent chez les bactéries . En médecine, ce qui contribue à la propagation de la la résistance aux antibiotiques, comme quand une bactérie acquiert des gènes de résistance, il peut rapidement les transférer à d'autres espèces. Le transfert horizontal de gènes des bactéries aux eucaryotes tels que la levure Saccharomyces cerevisiae et la fève dendroctone du adzuki Callosobruchus chinensis se est produite. Un exemple de transferts à grande échelle sont les eucaryote rotifères bdelloides, qui ont reçu une série de gènes de bactéries, les champignons et les plantes. Les virus peuvent également effectuer ADN entre organismes, permettant le transfert de gènes même à travers domaines biologiques.
Le transfert de gènes à grande échelle a eu lieu entre les ancêtres des cellules eucaryotes et les bactéries , au cours de l'acquisition de les chloroplastes et les mitochondries . Il est possible que les eucaryotes elles-mêmes proviennent de transferts horizontaux de gènes entre bactéries et archées.
Mécanismes
De Point de vue néo-darwinienne, l'évolution se produit quand il ya des changements dans la fréquence des allèles dans une population d'organismes métissage. Par exemple, l'allèle pour la couleur noire dans une population de papillons de plus en plus commun. Mécanismes qui peuvent conduire à des changements dans la fréquence des allèles comprennent la sélection naturelle , dérive génétique, auto-stop génétique, mutation et flux de gènes.
Sélection naturelle
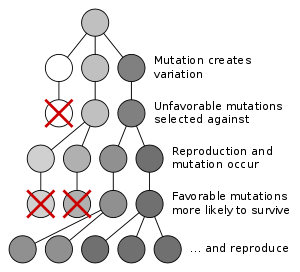
Evolution au moyen de la sélection naturelle est le processus par lequel les mutations génétiques qui améliorent la reproduction devenir et rester plus fréquente chez les générations successives d'une population. Il a souvent été appelé un mécanisme "de soi", car il découle nécessairement de trois faits simples:
- Variation héréditaire existe au sein de populations d'organismes.
- Organismes produisent plus de descendants que peut survivre.
- Ces descendants varient dans leur capacité à survivre et se reproduire.
Ces conditions produisent concurrence entre les organismes pour la survie et la reproduction. Par conséquent, les organismes avec des traits qui leur donnent un avantage sur leurs concurrents passent ces traits avantageux sur, tandis que les traits qui ne confèrent pas un avantage ne sont pas transmises à la génération suivante.
Le concept central de la sélection naturelle est le remise en forme évolutive d'un organisme. Fitness est mesurée par la capacité d'un organisme à survivre et se reproduire, ce qui détermine la taille de sa contribution génétique à la génération suivante. Cependant, remise en forme ne est pas le même que le nombre total de descendants: au lieu de remise en forme est indiqué par la proportion des générations suivantes qui portent les gènes d'un organisme. Par exemple, si un organisme pourrait bien survivre et se reproduire rapidement, mais sa progéniture étaient trop petits et faibles pour survivre, cet organisme ne aurait guère de contribution génétique aux générations futures et aurait donc faible fitness.
Si un allèle augmente remise en forme plus que les autres allèles de ce gène, puis avec chaque génération cet allèle deviendra plus commune au sein de la population. Ces traits sont dits être "sélectionné pour". Des exemples de traits qui peuvent améliorer la condition physique sont améliorées survie et augmenté la fécondité. Inversement, l'aptitude inférieure causé par des résultats d'allèles moins bénéfiques ou délétères dans cet allèle plus en plus rares - ils sont "sélectionné contre". Fait important, la remise en forme d'un allèle ne est pas une caractéristique fixe; si les changements de l'environnement, précédemment traits neutres ou nuisibles peuvent devenir traits bénéfiques et auparavant bénéfiques deviennent nuisibles. Cependant, même si la direction de sélection ne inverse de cette manière, les traits qui ont été perdus dans le passé peut ne pas ré-évoluer dans une forme identique (voir La loi de Dollo).
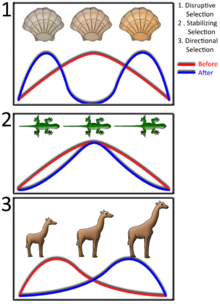
La sélection naturelle dans une population pour un trait qui peut varier dans une plage de valeurs, telles que la hauteur, peuvent être classés en trois types différents. Le premier est sélection directionnelle, ce qui est un changement dans la valeur moyenne d'un caractère au fil du temps - par exemple, les organismes lentement obtenir plus grand. Deuxièmement, sélection disruptive est la sélection des valeurs de traits extrêmes et se traduit souvent par deux valeurs différentes de devenir plus courante, avec la sélection contre la valeur moyenne. Ce serait quand organismes soit petit ou grand avaient un avantage, mais pas celles de taille moyenne. Enfin, dans stabilisation sélection il n'y a sélection par rapport aux valeurs extrêmes de traits sur les deux extrémités, ce qui provoque une diminution de la variance autour de la valeur moyenne et moins de diversité. Ce serait, par exemple, provoquer des organismes à devenir lentement la même hauteur.
Un cas particulier de la sélection naturelle est la sélection sexuelle, qui est la sélection pour un trait qui augmente succès d'accouplement en augmentant l'attractivité d'un organisme à des partenaires potentiels. Traits qui ont évolué par la sélection sexuelle sont particulièrement importants chez les mâles de certaines espèces animales, malgré des traits tels que bois, encombrants appels d'accouplement ou des couleurs vives qui attirent les prédateurs, diminuant la survie des hommes individuels. Ce désavantage de survie est compensée par le succès de reproduction plus élevé chez les hommes qui montrent ces difficile de faux, la sélection sexuelle traits.
La sélection naturelle fait le plus généralement la nature de la mesure contre laquelle les individus et les traits individuels, sont plus ou moins de chances de survivre. "Nature" en ce sens se réfère à un écosystème, à savoir un système dans lequel les organismes interagissent avec chaque autre élément, physique ainsi que biologique, dans leur locale environnement. Eugene Odum, un des fondateurs de l'écologie, un écosystème défini comme suit: «Toute unité qui comprend tous les organismes ... dans une zone donnée en interaction avec l'environnement physique de sorte qu'un flux d'énergie conduit à clairement défini la structure trophique, la diversité biotique et cycles des matières (ce est à dire: l'échange de matériel entre les parties vivantes et non vivantes) dans le système. " Chaque population dans un écosystème occupe une distincte niche, ou la position, avec des relations distinctes à d'autres parties du système. Ces relations impliquent l'histoire de la vie de l'organisme, sa position dans le chaîne alimentaire et de son aire de répartition géographique. Cette large compréhension de la nature permet aux scientifiques de délimiter forces spécifiques qui, ensemble, composent la sélection naturelle.
La sélection naturelle peut agir au différents niveaux d'organisation, tels que les gènes, des cellules, des organismes, des groupes d'organismes et d'espèces. La sélection peut agir à plusieurs niveaux simultanément. Un exemple de sélection se produisant au-dessous du niveau de l'organisme individuel sont appelés gènes transposons, qui peut se répliquer et se propager dans un génome. Sélection à un niveau supérieur à l'individu, telles que sélection de groupe, peut permettre l'évolution de la coopération, comme on le verra ci-dessous.
Mutation biaisée
En plus d'être une source importante de variante, la mutation peut également fonctionner comme un mécanisme de l'évolution quand il ya des probabilités différentes au niveau moléculaire pour différentes mutations se produisent, un processus connu sous le biais de mutation. Si deux génotypes, par exemple une avec le nucléotide G et un autre avec le nucléotide A dans la même position, le même remise en forme, mais la mutation de G à A arrive plus souvent que la mutation de A à G, alors génotypes A aura tendance à évoluer. Différent insertion vs biais suppression de mutation dans différents taxons peut conduire à l'évolution des différentes tailles de génome. Biais du développement ou de mutation ont également été observés dans évolution morphologique. Par exemple, selon le phénotype-première théorie de l'évolution, les mutations peut éventuellement provoquer la assimilation génétique de caractères qui étaient auparavant induite par l'environnement.
effets de polarisation de mutation sont superposées sur d'autres processus. Si la sélection favoriserait une ou l'autre de deux mutations, mais il n'y a pas d'avantage supplémentaire d'avoir à la fois, alors que la mutation se produit le plus fréquemment est celui qui est le plus susceptible de se fixer dans une population. Mutations conduisant à la perte de la fonction d'un gène sont beaucoup plus fréquentes que les mutations qui produisent un nouveau gène, pleinement fonctionnel. La plupart des pertes de mutations de fonction sont sélectionnés contre. Mais lorsque la sélection est faible, le biais de la mutation vers la perte de la fonction peut affecter l'évolution. Par exemple, les pigments ne sont plus utiles lorsque les animaux vivent dans l'obscurité de grottes, et ont tendance à se perdre. Ce type de perte de fonction peut se produire en raison d'un biais de mutation, et / ou parce que la fonction a un coût, et une fois l'avantage de la fonction a disparu, la sélection naturelle conduit à la perte. Perte de capacité de sporulation dans un bactérie cours de l'évolution de laboratoire semble avoir été causée par le biais de mutation, plutôt que la sélection naturelle contre le coût du maintien de la capacité de sporulation. Quand il n'y a pas de sélection pour la perte de la fonction, la vitesse à laquelle évolue la perte dépend de plus sur le taux de mutation qu'elle ne le fait sur le la taille effective de la population, ce qui indique qu'il est davantage motivé par des préjugés de mutation que par la dérive génétique.
La dérive génétique

La dérive génétique est le changement la fréquence des allèles d'une génération à la suivante se produit parce que les allèles sont soumis à L'erreur d'échantillonnage. Par conséquent, lorsque les forces sélectives sont absents ou relativement faible, les fréquences alléliques ont tendance à "dérive" vers le haut ou vers le bas de façon aléatoire (dans un marche aléatoire). Cette dérive se arrête quand un allèle devient finalement fixe, soit par disparaître de la population, ou de remplacer les autres allèles entièrement. La dérive génétique peut donc éliminer certains allèles d'une population attribuable au hasard seulement. Même en l'absence de forces sélectives, la dérive génétique peut provoquer deux populations distinctes qui ont commencé avec la même structure génétique à la dérive en deux populations divergentes avec différents ensembles d'allèles.
Il est généralement difficile de mesurer l'importance relative des processus de sélection et neutres, dont la dérive. L'importance comparative des forces adaptatifs et non adaptatifs dans la conduite de changement évolutif est un domaine de la recherche actuelle .
Le théorie neutre de l'évolution moléculaire proposé que la plupart des changements évolutifs sont le résultat de la fixation de mutations neutres par la dérive génétique. Ainsi, dans ce modèle, la plupart des changements génétiques dans une population sont le résultat de la pression constante mutation et la dérive génétique. Cette forme de la théorie neutre est désormais largement abandonnée, car elle ne semble pas correspondre à la variation génétique vu dans la nature. Cependant, une version plus récente et mieux prise en charge de ce modèle est la près la théorie neutre, où une mutation qui serait neutre dans une petite population ne est pas nécessairement neutre dans une grande population. D'autres théories alternatives proposent que la dérive génétique est éclipsé par d'autres forces stochastiques dans l'évolution, tels que auto-stop génétique, également connu comme projet de génétique.
Le temps pour un allèle neutre pour devenir fixé par la dérive génétique dépend de la taille de la population, avec fixation se produisant plus rapidement dans les petites populations. Le nombre d'individus dans une population ne est pas critique, mais au lieu d'une mesure connue sous le nom la taille effective de la population. La population effective est généralement plus petit que la population totale car elle prend en compte des facteurs tels que le niveau de consanguinité et l'étape du cycle de vie dans laquelle la population est la plus petite. La taille effective de la population ne peut pas être la même pour chaque gène dans la même population.
Auto-stop génétique
La recombinaison permet allèles sur le même brin d'ADN à se séparer. Cependant, le taux de recombinaison est faible (environ deux événements par chromosome par génération). En conséquence, les gènes rapprochées sur un chromosome peuvent pas toujours être mélangées l'une de l'autre et qui sont proches de gènes ont tendance à être hérités ensemble ensemble, un phénomène connu sous le nom tringlerie. Cette tendance est mesurée en trouvant comment souvent deux alleles apparaissent ensemble sur un seul chromosome par rapport à attentes, qui se appelle leur déséquilibre de liaison. Un ensemble d'allèles qui est habituellement hérité dans un groupe est appelé un haplotype. Cela peut être important quand un allèle dans un haplotype particulier est fortement bénéfique: la sélection naturelle peut conduire une balayage sélectif qui sera également provoquer les autres allèles dans l'haplotype à devenir plus fréquent dans la population; cet effet est appelé auto-stop génétique ou projet de génétique.Projet de génétique causé par le fait que certains gènes neutres sont génétiquement liés à d'autres qui sont sous sélection peut être partiellement capturé par une taille effective de la population appropriée.
Le flux de gènes
Le flux de gènes est l'échange de gènes entre les populations et entre espèces. La présence ou l'absence de flux de gènes change fondamentalement le cours de l'évolution. En raison de la complexité des organismes, des deux populations entièrement isolées finiront par évoluer incompatibilités génétiques à travers des processus neutres, comme dans le modèle Bateson-Dobzhansky-Muller, même si les deux populations restent essentiellement identiques en termes de leur adaptation à l'environnement.
Si la différenciation génétique entre les populations se développe, les flux de gènes entre les populations peut introduire des traits ou des allèles qui sont désavantageux dans la population locale, ce qui peut conduire à l'organisme au sein de ces populations à évoluer mécanismes qui empêchent l'accouplement avec des populations génétiquement éloignés, entraînant finalement l'apparition de nouvelles espèces. Ainsi, l'échange d'information génétique entre les individus est d'une importance fondamentale pour le développement du concept d'espèce biologique (BSC).
Lors de l'élaboration de la synthèse moderne, Sewall Wright a développé sa théorie de l'équilibre changeant que le flux génétique entre les populations partiellement isolées était un aspect important de l'évolution adaptative. Toutefois, récemment, il ya eu la critique substantielle de l'importance de la théorie de l'équilibre changeant.
Résultats
Evolution influence tous les aspects de la forme et le comportement des organismes. Les plus importantes sont les comportementales et physiques spécifiques adaptations qui sont le résultat de la sélection naturelle. Ces adaptations augmentent remise en forme en aidant des activités telles que la recherche de nourriture, éviter les prédateurs ou attirer les femelles. Les organismes peuvent également répondre à la sélection par coopérant avec l'autre, généralement en aidant leurs parents ou de se livrer mutuellement bénéfique en symbiose. À plus long terme, l'évolution produit de nouvelles espèces par les populations ancestrales fractionnement d'organismes dans de nouveaux groupes qui ne peuvent pas ou ne veulent pas se croiser.
Ces résultats de l'évolution sont parfois divisés en macroevolution, qui est l'évolution qui se produit au niveau ou au-dessus du niveau des espèces, telles que l'extinction et de spéciation et microévolution, qui est petits changements évolutifs, telles que des adaptations, au sein d'une espèce ou une population. En général, macroevolution est considéré comme le résultat de longues périodes de microévolution. Ainsi, la distinction entre micro et macro-évolution est pas un élément fondamental d'un - la différence est tout simplement le temps impliqué. Cependant, dans la macro-évolution, les traits des espèces entières peuvent être importants. Par exemple, une grande quantité de variation entre les individus d'une espèce permet d'adapter rapidement à de nouveaux habitats, ce qui réduit la chance de lui en voie d'extinction, tandis qu'un large éventail géographique augmente les chances de spéciation, en le rendant plus probable qu'une partie de la population sera devenir isolés. En ce sens, la microévolution et macroévolution pourrait impliquer la sélection à différents niveaux - avec microévolution agissant sur les gènes et les organismes, par rapport aux processus macroévolutifs tels que la sélection des espèces agissant sur des espèces entières et affectant leurs taux de spéciation et d'extinction.
Une idée fausse commune est que l'évolution a des objectifs ou des plans à long terme; réaliste toutefois, l'évolution n'a pas de but à long terme et ne produit pas nécessairement une plus grande complexité. Bien que espèces complexes ont évolué, ils se produisent comme un effet secondaire du nombre total des organismes de plus en plus et les formes simples de la vie demeurent plus fréquents dans la biosphère. Par exemple, l'immense majorité des espèces sont microscopiques procaryotes, qui forment environ la moitié du monde la biomasse malgré leur petite taille, et constituent la grande majorité de la biodiversité de la Terre. Organismes simples ont donc été la forme dominante de la vie sur Terre tout au long de son histoire et de continuer à être la principale forme de vie jusqu'à nos jours, avec la vie complexe ne comparaîtra plus diversifié, car il est plus perceptible. En effet, l'évolution des micro-organismes est particulièrement important de la recherche moderne de l'évolution , depuis leur reproduction rapide permet l'étude de l'évolution expérimentale et de l'observation de l'évolution et de l'adaptation en temps réel.
Adaptation
L'adaptation est le processus qui rend organismes mieux adapté à leur habitat. Aussi, l'adaptation à long terme peut se référer à un trait qui est important pour la survie de l'organisme. Par exemple, l'adaptation des dents de chevaux pour le broyage de l'herbe. En utilisant le terme d'adaptation pour le processus évolutif et adaptatif trait pour le produit (la partie ou une fonction corporelle), les deux sens du terme peuvent être distingués. Des adaptations sont produites par la sélection naturelle . Les définitions suivantes sont dues à Theodosius Dobzhansky.
- Adaptationest le processus évolutif par lequel un organisme devient de mieux en mesure de vivre dans sonhabitat ou habitats.
- Adaptabilitéest l'état d'être adapté: la mesure dans laquelle un organisme est capable de vivre et se reproduire dans un ensemble donné des habitats.
- Uncaractère adaptatifest un aspect du motif de développement de l'organisme qui permet ou améliore la probabilité de survie de cet organisme et de reproduction.
L'adaptation peut provoquer soit le gain d'une nouvelle fonctionnalité, ou la perte d'une fonction ancestrale. Un exemple montre que les deux types de modification est l'adaptation des bactéries aux antibiotiques sélection, et les variations génétiques provoquant une résistance aux antibiotiques à la fois par la modification de la cible du médicament, ou en augmentant l'activité de transporteurs qui pompent le médicament hors de la cellule. D'autres exemples frappants sont les bactéries Escherichia coli évolution de la capacité d'utiliser l'acide citrique en tant que nutriment dans une expérience de laboratoire à long terme, Flavobacterium évolue une nouvelle enzyme qui permet à ces bactéries de se développer sur les sous-produits de la fabrication de nylon, et la bactérie du sol Sphingobium évolution d'une toute nouvelle voie métabolique qui dégrade le synthétique pesticide pentachlorophénol. Une idée intéressante, mais encore controversée est que certaines adaptations pourraient augmenter la capacité des organismes à générer de la diversité génétique et adaptation par la sélection naturelle (organismes croissante de la capacité d'évolution).
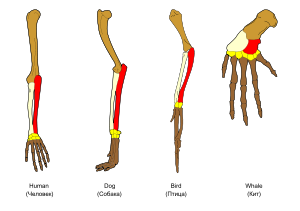
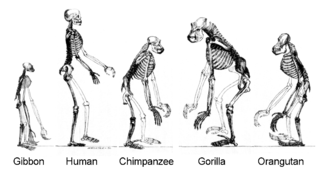
Adaptation se produit à travers la modification progressive des structures existantes. Par conséquent, les structures de l'organisation interne similaire peuvent avoir différentes fonctions dans des organismes apparentés. Ceci est le résultat d'une seule structure de ancestrale étant adapté pour fonctionner de différentes façons. Les os dans bat des ailes, par exemple, sont très semblables à celles de souris pieds et les mains de primates, en raison de la descente de toutes ces structures à partir d'un ancêtre commun des mammifères. Cependant, comme tous les organismes vivants sont liés dans une certaine mesure, même des organes qui semblent avoir peu ou pas de similitude de structure, tels que les arthropodes , les calmars et les yeux, vertébrés ou les membres et les ailes d'arthropodes et les vertébrés, peuvent compter sur un ensemble commun de gènes homologues qui contrôlent leur assemblage et la fonction; cela se appelle homologie profonde.
Au cours de l'évolution, certaines structures peuvent perdre leur fonction d'origine et devenir structures rudimentaires . Ces structures peuvent avoir peu ou pas de fonction dans une espèce de courant, mais ont une fonction claire des espèces ancestrales, ou d'autres espèces étroitement liées. Des exemples comprennent pseudogènes, les restes non-fonctionnels d'yeux dans les poissons cavernicoles aveugles, des ailes chez les oiseaux de voler, et la présence d'os de la hanche chez les baleines et les serpents. Des exemples de structures rudimentaires chez les humains comprennent des dents de sagesse, le coccyx, l' annexe vermiforme, et autres vestiges de comportement tels que la chair de poule et les réflexes primitifs.
Cependant, de nombreux traits qui semblent être de simples adaptations sont en fait exaptations: structures adaptées à l'origine pour une fonction, mais qui par hasard sont devenus quelque peu utile pour une autre fonction dans le processus. Un exemple est le lézard africaine holaspis guentheri , qui a développé une tête extrêmement plat pour cacher dans les crevasses, comme on peut le voir en regardant ses proches parents. Cependant, dans cette espèce, la tête est devenue tellement aplati qu'il aide à glissement d'arbre en arbre-un exaptation. Dans les cellules, des machines moléculaires tels que l'bactérienne flagelles et machines de tri des protéines ont évolué par le recrutement de plusieurs protéines préexistantes qui avaient auparavant des fonctions différentes. Un autre exemple est le recrutement d'enzymes de la glycolyse et métabolisme des xénobiotiques pour servir de protéines structurelles appelées cristallines dans les lentilles de de organismes les yeux .
Un principe essentiel de l'écologie est celle de l'exclusion compétitive: deux espèces ne peuvent occuper le même créneau dans le même environnement pendant une longue période. Par conséquent, la sélection naturelle a tendance à forcer les espèces à s'adapter à différents niches écologiques. Cela peut signifier que, par exemple, deux espèces de cichlidés adapter à vivre dans différents habitats, qui permettra de minimiser la concurrence entre eux pour la nourriture.
Une zone d'enquête en cours dans la biologie évolutive du développement est la base du développement d'adaptations et de exaptations. Cette recherche porte sur l'origine et l'évolution du développement embryonnaire et comment des modifications de développement et les processus de développement produisent de nouvelles caractéristiques. Ces études ont montré que l'évolution peut altérer le développement pour produire de nouvelles structures, telles que les structures osseuses embryonnaires qui se développent dans la mâchoire à la place d'autres animaux qui font partie de l'oreille moyenne chez les mammifères. Il est également possible pour les structures qui ont été perdus dans l'évolution de réapparaître en raison de changements dans les gènes du développement, comme une mutation dans poulets provoquant des embryons de se développer dents semblables à celles des crocodiles . Il est désormais clair que la plupart des modifications dans la forme d'organismes sont dues à des changements dans un petit ensemble de gènes conservés.
Co-évolution

Les interactions entre les organismes peuvent produire à la fois des conflits et de la coopération. Lorsque l'interaction entre paires d'espèces, comme un agent pathogène et un hôte, ou d'un prédateur et sa proie, ces espèces peuvent développer des séries d'adaptations adaptées. Ici, l'évolution d'une espèce provoque des adaptations dans une deuxième espèce. Ces changements dans la seconde espèce, puis, à leur tour, provoquent de nouvelles adaptations de la première espèce. Ce cycle de sélection et la réponse est appelé co-évolution. Un exemple est la production de la tétrodotoxine dans le triton à peau rugueuse et l'évolution de la résistance de la tétrodotoxine dans son prédateur, la couleuvre rayée. Dans cette paire de prédateur-proie, une course aux armements évolution a produit des niveaux élevés de la toxine dans le triton et des niveaux élevés de résistance correspondante de toxine dans le serpent.
Coopération
Toutes les interactions co-évolué entre les espèces comportent pas de conflit. De nombreux cas d'interactions mutuellement bénéfiques ont évolué. Par exemple, une coopération extrême existe entre les plantes et les champignons mycorhiziens qui poussent sur leurs racines et de l'aide la plante à absorber les nutriments du sol. C'est un relation réciproque que les plantes fournissent les champignons avec des sucres de la photosynthèse. Ici, les champignons poussent réellement à l'intérieur des cellules végétales, leur permettant d'échanger des éléments nutritifs avec leurs hôtes, tout en envoyant des signaux qui suppriment l'usine système immunitaire .
Coalitions entre les organismes de la même espèce ont également évolué. Un cas extrême est la eusociality trouve dans les insectes sociaux, tels que les abeilles , les termites et les fourmis , où les insectes stériles nourrissent et gardent le petit nombre d'organismes dans une colonie qui sont capables de se reproduire. Sur une échelle encore plus réduite, les cellules somatiques qui constituent le corps d'un animal limitent leur reproduction afin qu'ils puissent maintenir un organisme stable, qui supporte alors un petit nombre de de l'animal les cellules germinales de produire une descendance. Ici, les cellules somatiques réagissent aux signaux spécifiques qui les instruisent si de croître, restent comme ils sont, ou qui meurent. Si les cellules ignorent ces signaux et se multiplient de façon inappropriée, leur croissance incontrôlée provoque le cancer.
Cette coopération au sein des espèces peut avoir évolué à travers le processus de sélection de parentèle, qui est l'endroit où un organisme agit pour aider à élever la progéniture d'un parent. Cette activité est sélectionné pour parce que si l' aide individuelle contient allèles qui favorisent l'activité aidant, il est probable que sa famille sera également contenir ces allèles et donc ces allèles sera transmis. D'autres procédés qui peuvent promouvoir la coopération comprennent la sélection de groupe, où la coopération offre des avantages à un groupe d'organismes.
Spéciation
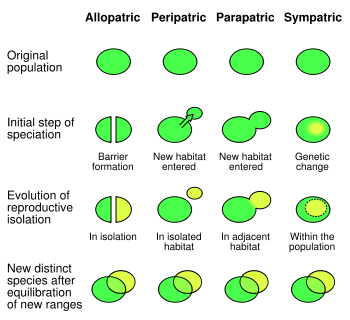
Spéciation est le processus par lequel une espèce diverge en deux ou plusieurs espèces descendantes.
Il ya de multiples façons de définir le concept de «espèce». Le choix de la définition dépend des particularités des espèces concernées. Par exemple, certains concepts d'espèces appliquent plus facilement vers les organismes sexuellement reproduire tandis que d'autres se prêtent mieux vers les organismes asexués. Malgré la diversité des différents concepts d'espèces, ces différents concepts peuvent être placés dans l'une des trois grandes approches philosophiques: métissage, écologique et phylogénétique. Le concept d'espèce biologique (BSC) est un exemple classique de l'approche de métissage. Défini par Ernst Mayr en 1942, le BSC affirme que «les espèces sont des groupes de réellement ou potentiellement métissage des populations naturelles, qui sont la reproduction isolés à partir d'autres groupes". Malgré son utilisation large et à long terme, le BSC comme les autres est pas sans controverse, par exemple parce que ces concepts ne peuvent pas être appliquées aux procaryotes, et ce qu'on appelle le problème des espèces. Certains chercheurs ont tenté une définition moniste fédérateur des espèces, tandis que d'autres adoptent une approche pluraliste et suggèrent qu'il peut y avoir différentes façons d'interpréter logiquement la définition d'une espèce. "
Les obstacles à la reproduction entre deux populations divergentes sexuelles sont nécessaires pour les populations à devenir de nouvelles espèces. Le flux de gènes peut ralentir ce processus, en écartant les nouvelles variantes génétiques aussi pour les autres populations. Selon la distance deux espèces ont divergé depuis leur ancêtre commun le plus récent, il peut toujours être possible pour eux de produire une descendance, comme avec les chevaux et les ânes accouplement pour produire des mules. Tel hybrides sont généralement infertile. Dans ce cas, les espèces étroitement liées peuvent régulièrement se croisent, mais les hybrides seront sélectionnés contre et les espèces resteront distincts. Cependant, des hybrides viables sont parfois formé et ces nouvelles espèces peuvent soit avoir des propriétés intermédiaires entre leurs espèces parentes, ou de posséder un tout nouveau phénotype. L'importance de l'hybridation dans la production de nouvelles espèces d'animaux est difficile, bien que des cas ont été observés dans de nombreux types d'animaux, avec la rainette versicolore est un exemple particulièrement bien étudié.
Spéciation a été observée plusieurs fois dans les deux conditions de laboratoire contrôlées et dans la nature. Dans les organismes à reproduction sexuée, les résultats de la spéciation de l'isolement reproductif suivis par la divergence généalogiques. Il ya quatre mécanismes de spéciation. Le plus commun chez les animaux est spéciation allopatrique, qui se produit dans les populations isolées géographiquement abord, comme par fragmentation ou la migration habitat. Sélection dans ces conditions, peut produire des changements très rapides dans l'apparence et le comportement des organismes. Comme la sélection et la dérive agissent indépendamment sur les populations isolées du reste de leur espèce, la séparation peut éventuellement produire des organismes qui ne peuvent pas se croiser.
Le second mécanisme de spéciation est spéciation péripatrique, qui se produit lorsque de petites populations d'organismes deviennent isolés dans un nouvel environnement. Cela diffère de la spéciation allopatrique en ce que les populations isolées sont numériquement beaucoup plus petite que la population parentale. Ici, l' effet fondateur provoque spéciation rapide après une augmentation de la consanguinité augmente sélection sur homozygotes, menant à un changement génétique rapide.
Le troisième mécanisme de spéciation est spéciation parapatriques. Ceci est similaire à péripatrique spéciation en ce qu'une petite population entre dans un nouvel habitat, mais diffère en ce que il n'y a pas de séparation physique entre ces deux populations. Au lieu de cela, la spéciation résulte de l'évolution des mécanismes qui réduisent le flux génétique entre les deux populations. En général, cela se produit quand il ya eu un changement radical dans l'environnement au sein de l'habitat des espèces parentales. Un exemple est l'herbe flouve odorante , qui peut subir la spéciation parapatriques en réponse à la pollution métallique localisée des mines. Ici, les plantes évoluer qui ont une résistance à des niveaux élevés de métaux dans le sol. Sélection contre le croisement avec la population parentale sensible métal produit un changement progressif dans le temps de la floraison des plantes résistantes métalliques, qui a finalement produites isolement reproducteur complet. Sélection contre des hybrides entre les deux populations peut provoquer renfort , qui est l'évolution des traits qui favorisent l'accouplement sein d'une espèce, ainsi que le déplacement de caractère, ce qui est quand deux espèces deviennent plus distinctes en apparence.
Enfin, dans espèces de spéciation sympatrique divergent sans isolement géographique ou de changements dans l'habitat. Cette forme est rare car même une petite quantité de flux de gènes peut supprimer les différences génétiques entre les parties d'une population. Généralement, spéciation sympatrique chez les animaux nécessite l'évolution des deux différences génétiques et accouplement non aléatoire, pour permettre l'isolement reproductif d'évoluer.
Un type de spéciation sympatrique implique le croisement de deux espèces apparentées pour produire une nouvelle espèce hybride. Ce ne sont pas fréquentes chez les animaux comme les hybrides d'animaux sont généralement stérile. Ceci est parce que pendant la méiose les chromosomes homologues de chaque parent sont de différentes espèces et ne peuvent jumeler avec succès. Cependant, il est plus fréquent dans les usines parce que les plantes doublent souvent leur nombre de chromosomes, pour former polyploïdes. Cela permet aux chromosomes de chacune des espèces parentales pour former des paires lors de la méiose, puisque les chromosomes de chaque parent sont représentés par une paire déjà. Un exemple d'un tel événement de spéciation est où l'espèce de la plante Arabidopsis thaliana et Arabidopsis arenosa Croisée de donner les nouvelles espèces Arabidopsis suecica . Ce qui est arrivé il ya environ 20.000 ans, et le processus de spéciation a été répété dans le laboratoire, qui permet l'étude des mécanismes génétiques impliqués dans ce processus. En effet, un doublement des chromosomes d'une espèce peut être une cause fréquente de l'isolement reproductif, comme la moitié des chromosomes doublé seront inégalée lors de la reproduction des organismes undoubled.
évènements de spéciation sont importants dans la théorie de l'équilibre ponctué , qui représente la tendance dans le registre fossile de courts "rafales" d'évolution entrecoupées de périodes relativement longues de stase, où les espèces restent relativement stables. Dans cette théorie, la spéciation et évolution rapide sont liés, avec la sélection naturelle et la dérive génétique agissant plus fortement sur les organismes soumis à la spéciation dans de nouveaux habitats ou de petites populations. En conséquence, les périodes de stagnation dans les archives fossiles correspondent à la population parentale et les organismes subissant spéciation et évolution rapide se trouvent dans de petites populations ou les habitats limités géographiquement et donc rarement préservés sous forme de fossiles.
Extinction

Extinction est la disparition d'une espèce entière. L'extinction est pas un événement inhabituel, comme des espèces apparaissent régulièrement dans la spéciation et disparaissent grâce à l'extinction. Presque toutes les espèces animales et végétales qui ont vécu sur la Terre sont maintenant éteintes, et l'extinction semble être le sort ultime de toutes les espèces. Ces extinctions se sont produites en continu tout au long de l'histoire de la vie, bien que le taux de pointes d'extinction en masse occasionnels événements d'extinction. Le extinction crétacé-tertiaire, au cours de laquelle les dinosaures non-aviaires se sont éteints, est le plus connu, mais le plus tôt Permien événement d'extinction -Triassic était encore plus sévère, avec environ 96% des espèces en situation d'extinction. Le événement d'extinction de l'Holocène est une extinction de masse continue associée à l'expansion de l'humanité à travers le monde au cours des mille dernières années. Les taux d'extinction actuels sont 100-1000 fois plus élevé que le taux de base et jusqu'à 30% des espèces actuelles peut être éteinte par le mi 21ème siècle. Les activités humaines sont maintenant la principale cause de l'événement d'extinction en cours; le réchauffement climatique peut encore accélérer à l'avenir.
Le rôle de l'extinction de l'évolution est pas très bien compris et peut dépendre du type d'extinction est considéré. Les causes des événements d'extinction continues "bas niveau", qui forment la majorité des extinctions, peuvent être le résultat de la compétition entre les espèces pour des ressources limitées ( d'exclusion compétitive). Si une espèce peuvent supplanter l'autre, ce qui pourrait produire de la sélection des espèces, avec les espèces monteur survivant et les autres espèces étant conduit à l'extinction. Les extinctions de masse intermittents sont également importants, mais au lieu d'agir comme une force sélective, ils réduisent considérablement la diversité d'une manière non spécifique et de promouvoir des éclats de rapide évolution et de la spéciation chez les survivants.
Histoire de l'évolution de la vie
Origine de la vie
La chimie hautement énergétique est pensé pour avoir produit une molécule d'auto-réplication autour 4000000000 années auparavant, et un demi-milliard d'années plus tard, le dernier ancêtre commun de toute vie existait. Le courant consensus scientifique est que le complexe de la biochimie qui fait la vie venait de réactions chimiques simples. Le début de la vie peut avoir inclus des molécules d'auto-réplication, tels que l'ARN et l'assemblage de cellules simples.
Ascendance commune
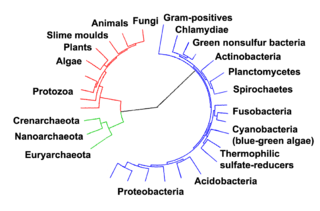
Tous les organismes sur la Terre sont les descendants d'un ancêtre commun ou la piscine de gène ancestral. Espèces actuelles sont une étape dans le processus d'évolution, avec leur diversité le produit d'une longue série de spéciation et d'extinction des événements. Le descendance commune des organismes a été déduite de quatre faits simples sur les organismes: d'abord, ils ont des distributions géographiques qui ne peuvent pas être expliquées par l'adaptation locale. Deuxièmement, la diversité de la vie est pas un ensemble d'organismes complètement uniques, mais les organismes qui partagent des similitudes morphologiques. Troisièmement, les traits rudimentaires sans objectif clair ressemblent traits ancestraux fonctionnels et enfin, que les organismes peuvent être classés en utilisant ces similitudes dans une hiérarchie de groupes imbriqués - semblable à un arbre généalogique. Cependant, la recherche moderne a suggéré que, en raison du transfert horizontal de gènes, cette « arbre de vie »peut être plus compliqué que d'un arbre de branchement simple car certains gènes se sont propagées indépendamment entre espèces apparentées de loin.
Espèces passées ont également laissé des enregistrements de leur histoire évolutive. fossiles , avec l'anatomie comparée des organismes actuels, constituent le morphologique, ou anatomique, fiche. En comparant les anatomies des deux espèces modernes et éteints, les paléontologues peuvent déduire les lignées de ces espèces. Cependant, cette approche est plus efficace pour les organismes qui avaient parties dures, comme des coquillages, des os ou les dents. En outre, comme les procaryotes tels que les bactéries et les archées partager un ensemble limité de morphologies communs, leurs fossiles ne fournissent pas d'informations sur leur ascendance.
Plus récemment, la preuve de la descendance commune est venue de l'étude de biochimiques similitudes entre les organismes. Par exemple, toutes les cellules vivantes utilisent le même ensemble de base de nucléotides et des acides aminés . Le développement de génétique moléculaire a révélé le record de l'évolution à gauche de organismes des génomes: Les rencontres lorsque les espèces ont divergé par l' horloge moléculaire produite par des mutations. Par exemple, ces comparaisons de séquences d'ADN ont révélé que les humains et les chimpanzés partagent 98% de leurs génomes et d'analyser les quelques domaines où ils diffèrent aide la lumière de hangar quand l'ancêtre commun de ces espèces existaient.
Évolution de la vie
Procaryotes habité la Terre d'environ 3-4 milliards d'années il ya. Pas de changements évidents dans la morphologie ou organisation cellulaire ont eu lieu dans ces organismes au cours des milliards d'années. Les cellules eucaryotes sont apparues entre 1,6 - Il ya 2,7 milliards années. Le prochain changement majeur dans la structure cellulaire est venu lorsque les bactéries ont été engloutis par les cellules eucaryotes, dans une association coopérative appelé endosymbiose. Les bactéries englouti et la cellule hôte ont ensuite subi une co-évolution, avec les bactéries évoluant en soit mitochondries ou hydrogénosomes. Un autre engloutissement des organismes de cyanobactéries comme a conduit à la formation de chloroplastes dans les algues et les plantes.
L'histoire de la vie était celui des eucaryotes unicellulaires procaryotes et les archées, jusqu'à il ya environ 610 millions d'années quand les organismes multicellulaires ont commencé à apparaître dans les océans dans lapériode d'Ediacara. Le évolution de multicellularité eu lieu dans plusieurs événements indépendants, dans des organismes aussi divers que éponges, les algues brunes , cyanobactéries, moules et de bouemyxobactéries.
Peu de temps après l'apparition de ces premiers organismes multicellulaires, une quantité remarquable de la diversité biologique est apparu sur environ 10 millions d'années, dans un événement appelé l' explosion cambrienne . Ici, la majorité des types d'animaux modernes apparu dans les archives fossiles, ainsi que des lignées uniques qui ont disparu par la suite. Différents déclencheurs pour l'explosion cambrienne ont été proposés, y compris l'accumulation de l'oxygène dans l' atmosphère à partir de la photosynthèse .
Il ya environ 500 millions d'années, les plantes et les champignons ont colonisé la terre et furent bientôt suivis par les arthropodes et autres animaux. Les insectes ont particulièrement bien réussi et aujourd'hui encore, constituent la majorité des espèces animales. Amphibiens abord apparus il ya environ 364.000.000 années, suivie par les premiers amniotes et les oiseaux , il ya environ 155 millions d'années (à la fois de " reptile "-comme lignées), les mammifères il ya environ 129.000.000 années, Homininae il ya environ 10 millions d'années et l'homme moderne il ya environ 250000 années. Cependant, en dépit de l'évolution de ces grands animaux, les petits organismes semblables aux types qui ont évolué au début de ce processus continuent d'être très réussie et de dominer la Terre, avec la majorité des deux espèces étant la biomasse et les procaryotes.
Applications
Concepts et modèles utilisés dans la biologie évolutionniste, comme la sélection naturelle, ont de nombreuses applications.
Sélection artificielle est la sélection intentionnelle de traits dans une population d'organismes. Cela a été utilisé depuis des milliers d'années dans la domestication des plantes et des animaux. Plus récemment, une telle sélection est devenue une partie essentielle de l'ingénierie génétique, avec des marqueurs sélectionnables tels que les gènes de résistance aux antibiotiques utilisés pour manipuler l'ADN. Dans des cycles répétés de mutation et de sélection des protéines avec des propriétés intéressantes ont évolué, par exemple modifiée enzymes et de nouveaux anticorps , dans un processus appelé l'évolution dirigée.
Comprendre les changements qui ont eu lieu au cours de l'évolution de l'organisme peut révéler les gènes nécessaires à la construction des parties du corps, des gènes qui peuvent être impliqués dans les droits de troubles génétiques. Par exemple, le tétra mexicain est un albinos cavefish qui a perdu son acuité visuelle cours de l'évolution. Elevage ensemble différentes populations de ce poisson aveugle produites certains descendants avec les yeux fonctionnels, puisque les différentes mutations avaient eu lieu dans les populations isolées qui avaient évolué dans différentes grottes. Cela a aidé à identifier les gènes nécessaires à la vision et la pigmentation.
En informatique , des simulations d'évolution en utilisant les algorithmes évolutionnaires et la vie artificielle ont commencé dans les années 1960 et a été étendu à la simulation de la sélection artificielle. évolution artificielle est devenue une méthode d'optimisation largement reconnu comme un résultat du travail d' Ingo Rechenberg dans les années 1960. Il a utilisé stratégies d'évolution de résoudre les problèmes d'ingénierie complexes. Les algorithmes génétiques en particulier est devenu populaire à travers l'écriture de John Holland. Les applications pratiques comprennent également l'évolution automatique de programmes informatiques. Les algorithmes évolutionnaires sont maintenant utilisés pour résoudre des problèmes multidimensionnels plus efficace que les logiciels produits par les concepteurs de l'homme et aussi pour optimiser la conception des systèmes.
Réponses sociales et culturelles

Au 19e siècle, en particulier après la publication de L'Origine des Espèces en 1859, l'idée que la vie avait évolué était une source active de débat académique centrée sur les implications philosophiques, sociales et religieuses de l'évolution. Aujourd'hui, la théorie synthétique de l'évolution est acceptée par une grande majorité des scientifiques. Cependant, l'évolution reste un concept controversé pour certains théistes.
Tandis que diverses religions et confessions se sont réconciliés avec leurs croyances évolution à travers des concepts tels que l'évolution théiste, il ya des créationnistes qui croient que l'évolution est contredite par les mythes de la création trouvés dans leurs religions et qui soulève diverses objections à l'évolution. Comme il a été démontré par les réponses à la publication des Vestiges de l'histoire naturelle de la création en 1844, l'aspect le plus controversé de la biologie évolutionniste est l'implication de l'évolution humaine que les humains partagent un ancêtre commun avec les singes et que les facultés mentales et morales de l'humanité ont les mêmes types de causes naturelles que les autres traits hérités chez les animaux. Dans certains pays, notamment les États-Unis, ces tensions entre la science et la religion ont alimenté le courant controverse création-évolution, un conflit religieux en se concentrant sur la politique et l'éducation du public. si d'autres domaines scientifiques tels que la cosmologie et de sciences de la Terre aussi en conflit avec des interprétations littérales de nombreux textes religieux , des expériences de biologie de l'évolution nettement plus l'opposition des littéralistes religieuses.
L'enseignement de l'évolution dans les classes de biologie de l'enseignement secondaire américains était rare dans la plupart de la première moitié du 20e siècle. Le décision procès Scopes de 1925 a causé le sujet à devenir très rare en Amérique manuels de biologie secondaires pour une génération, mais il a été progressivement ré introduit sur une génération plus tard et juridiquement protégé avec 1968, le Epperson v. Arkansas décision. Depuis lors, la croyance religieuse concurrente du créationnisme a été légalement refusé dans les programmes des écoles secondaires dans diverses décisions dans les années 1970 et 1980, mais il est revenu en forme pseudoscientifique que la conception intelligente, d'être exclus une fois de plus dans le 2005 Kitzmiller c. Dover District School Area cas.