

Fongus
Saviez-vous ...
Cette sélection de wikipedia a été choisi par des bénévoles aidant les enfants SOS de Wikipedia pour cette sélection Wikipedia pour les écoles. Cliquez ici pour en savoir plus sur le parrainage d'enfants.
| Champignons Plage temporelle: début du Dévonien -Recent (mais voir le texte ) | |
|---|---|
 | |
| À droite en haut à gauche: Amanita muscaria, un basidiomycète; Sarcoscypha coccinea, un ascomycète; pain recouvert de moisissures ; une chytride; une Aspergillus conidiophore. | |
| Classification scientifique | |
| Domaine: | Eucaryotes |
| (Non classé): | Opisthokonta |
| Uni: | Champignons ( L. , 1753) RT Moore, 1980 |
| Embranchements / Phyla / embranchements | |
Dikarya (inc. Deuteromycota)
Embranchements incertae sedis
| |
Un champignon (pron .: / fa ʌ ŋ ɡ ə s /; pluriel: des champignons ou des champignons) est membre d'un grand groupe d' eucaryotes organismes qui comprend des micro-organismes tels que les levures et les moisissures ( L'anglais britannique: moules), ainsi que les plus familiers des champignons . Ces organismes sont classés en tant que royaume, des champignons, qui est séparée de plantes , les animaux et les bactéries . Une différence majeure est que les cellules fongiques ont les parois des cellules qui contiennent la chitine, contrairement aux parois cellulaires de plantes, qui contiennent cellulose. Ceux-ci et d'autres différences montrent que les champignons constituent un seul groupe d'organismes apparentés, nommé le Eumycota (champignons vrais ou Eumycètes), qui part d'un ancêtre commun (un groupe monophylétique). Ce groupe fongique est distincte de la structure semblable myxomycètes (myxomycètes) et oomycètes (moules d'eau). La discipline de la biologie consacrée à l'étude des champignons est connu comme mycologie (du grec μύκης, Mukes, qui signifie «champignon»). Mycologie a souvent été considérée comme une branche de la botanique , même si ce est un royaume distinct biologique taxonomie. Des études génétiques ont montré que les champignons sont plus étroitement liées aux animaux que pour les plantes.
Abondant dans le monde entier, la plupart des champignons sont discrètes en raison de la petite taille de leurs structures et de leur modes de vie dans le sol, cryptiques sur la matière morte, et que symbiotes des végétaux, des animaux, ou d'autres champignons. Ils peuvent devenir visible lorsque fructification, soit comme des champignons ou des moisissures. Les champignons jouent un rôle essentiel dans la décomposition de la matière organique et des rôles fondamentaux dans cycle des éléments nutritifs et l'échange. Ils ont longtemps été utilisés comme une source directe de nourriture, tels que les champignons et truffes, comme un levain agent pour pain, et dans fermentation de divers produits alimentaires tels que le vin , la bière , et la sauce de soja. Depuis les années 1940, des champignons ont été utilisées pour la production d' antibiotiques , et, plus récemment, divers enzymes produites par des champignons sont utilisés industriellement et en détergents. Les champignons sont également utilisés comme pesticides biologiques pour contrôler les mauvaises herbes, les maladies des plantes et les insectes nuisibles. De nombreuses espèces produisent composés bioactifs appelés mycotoxines, comme alcaloïdes et polycétides, qui sont toxiques pour les animaux y compris l'homme. Les structures de fructification de quelques espèces contiennent composés psychotropes et sont consommés récréative ou traditionnelle des cérémonies spirituelles. Les champignons peuvent décomposer les matériaux et bâtiments manufacturés, et devenir significative des agents pathogènes humains et d'autres animaux. Les pertes de récoltes dues aux maladies fongiques (par exemple pyriculariose du riz de la maladie) ou de la nourriture altération peut avoir un grand impact sur l'homme l'approvisionnement alimentaire et les économies locales.
Le royaume de champignon englobe une énorme diversité de taxons avec écologies variées, stratégies de cycle de vie, et morphologies allant de aquatique unicellulaire chytrides à gros champignons. Cependant, on sait peu de la véritable biodiversité des champignons Uni, qui a été estimée à 1500000 à 5000000 espèces, avec environ 5% d'entre eux ayant été officiellement classés. Depuis le pionnier 18ème et 19ème siècle travaux taxonomiques de Carl Linnaeus , Christian Hendrik Persoon, et Elias Magnus Fries, les champignons ont été classés en fonction de leur morphologie (par exemple, des caractéristiques telles que la couleur des spores ou des caractéristiques microscopiques) ou physiologie. Les progrès de la génétique moléculaire ont ouvert la voie à analyse de l'ADN à intégrer dans la taxonomie, qui a parfois contesté les groupements historiques basés sur la morphologie et d'autres traits. Les études phylogénétiques publiés dans la dernière décennie ont contribué à remodeler la classification des champignons Uni, qui est divisé en une sous-règne, sept phylums, et dix sous-embranchements.
Étymologie
Le mot champignon anglais est adopté directement à partir de la latine champignon (champignon), utilisé dans les écrits de Horace et Pline. Ce à son tour est dérivé du grec mot sphongos / σφογγος («éponge»), qui se réfère à la structures macroscopiques et la morphologie des champignons et des moisissures; la racine est également utilisé dans d'autres langues, comme l'allemand Schwamm ("éponge") et Schimmel («moule»). L'utilisation du mot mycologie, qui est dérivé du grec mykes / μύκης (champignons) et les logos / λόγος (discours), pour désigner l'étude scientifique des champignons est probablement originaire en 1836 avec naturaliste anglais La publication de miles Joseph Berkeley La Flora anglaise de Sir James Edward Smith, Vol. 5.
Caractéristiques

1- 2- hyphes mur septum 3- 4- mitochondrie Vacuole 5- cristal Ergostérol 6- ribosome 7- 8- réticulum endoplasmique Nucleus corps 9- 10- membrane lipidique plasma 11- Appareil Spitzenkörper 12 Golgi
Avant l'introduction de méthodes moléculaires pour l'analyse phylogénétique, taxonomistes considérés comme des champignons d'être membres du règne végétal en raison de similitudes dans le mode de vie: les deux champignons et les plantes sont principalement immobile, et présentent des similitudes dans la morphologie générale et de l'habitat de croissance. Comme les plantes, les champignons se développent souvent dans le sol, et dans le cas des champignons forment remarquable fructifications, qui portent parfois ressemblance avec des plantes telles que les mousses . Les champignons sont maintenant considérés comme un royaume séparé, distinct de plantes et d'animaux, dont ils semblent avoir divergé il ya environ un milliard d'années. Certaines caractéristiques morphologiques, biochimiques et génétiques sont partagés avec d'autres organismes, tandis que d'autres sont uniques aux champignons, en les séparant clairement des autres royaumes:
Caractéristiques communes:
- Avec d'autres eucaryotes : Comme d'autres eucaryotes, les cellules fongiques contiennent membranaires noyaux avec chromosomes qui contiennent de l'ADN avec régions non codantes appelées des introns et des régions codantes appelées exons. En outre, les champignons possèdent cytoplasmique membranaire organites tels que les mitochondries , stérol contenant des membranes, et ribosomes du Type 80S. Ils ont une gamme caractéristique de glucides solubles et en composés de stockage, y compris alcools de sucre (par exemple, mannitol), les disaccharides (par exemple, trehalose), et des polysaccharides (par exemple, glycogène, qui est également constaté chez les animaux).
- Chez les animaux: le manque champignons chloroplastes et sont organismes hétérotrophes, nécessitant préformés composés organiques comme source d'énergie.
- Avec les plantes: Fungi possèdent une paroi cellulaire et vacuoles. Ils se reproduisent par des moyens tant sexuée et asexuée, et comme groupes de plantes de base (telles que les fougères et les mousses ) produisent spores. Comme pour les mousses et les algues, les champignons ont typiquement noyaux haploïdes.
- Avec euglénoïdes et les bactéries: Les champignons supérieurs, euglénoïdes, et certaines bactéries produisent l' acide aminé L-lysine en particulier étapes de la biosynthèse, appelé le α-aminoadipate voie.
- Les cellules de la plupart des champignons poussent comme tubulaire, allongée, et les structures filiformes (filamenteux) et sont appelés hyphes, qui peut contenir plusieurs noyaux et d'étendre à leurs extrémités. Chaque conseil contient un ensemble de agrégée des vésicules cellulaires, comprenant des structures des protéines , des lipides et d'autres molécules organiques, appelés Spitzenkörper. Les deux champignons et oomycètes croître comme les cellules des hyphes filamenteux. En revanche, les organismes, tels que filamenteuse aspect semblable les algues vertes, se développent par division cellulaire répétée au sein d'une chaîne de cellules.
- En commun avec certaines espèces de plantes et d'animaux, et plus de 60 espèces fongiques présentent le phénomène de bioluminescence.
Caractéristiques uniques:
- Certaines espèces poussent comme des levures unicellulaires qui se reproduisent par bourgeonnement ou fission binaire. Champignons dimorphes peuvent basculer entre une phase et une phase levure fongique, en réponse aux conditions environnementales.
- La paroi cellulaire fongique est composé de glucanes et chitine; tandis que les premiers composés sont également présents dans les plantes et cette dernière dans la exosquelette des arthropodes , des champignons sont les seuls organismes qui combinent ces deux molécules structurales dans leur paroi cellulaire. En revanche pour les plantes et les oomycètes, les parois cellulaires des champignons ne contiennent pas de cellulose.

La plupart des champignons ne ont pas un système efficace pour le transport longue distance de l'eau et des nutriments, tels que le xylème et phloème dans de nombreuses plantes. Pour surmonter ces limitations, des champignons, tels que Armillaria, forme rhizomorphes, qui ressemblent et exécuter des fonctions similaires à la racines de plantes. Une autre caractéristique commune de plantes comprend un voie de biosynthèse pour la production de terpènes qui utilise acide mévalonique et pyrophosphate comme blocs de construction chimiques. Cependant, les plantes ont une voie de terpène supplémentaire dans leurs chloroplastes une structure champignons ne possèdent pas. Les champignons produisent plusieurs des métabolites secondaires qui sont similaires ou identiques à celles de la structure faite par les plantes. Un grand nombre des enzymes végétales et fongiques qui rendent ces composés diffèrent l'une de l'autre dans séquence et d'autres caractéristiques, ce qui indique origines distinctes et l'évolution de ces enzymes dans les champignons et les plantes.
Diversité

Les champignons ont une distribution dans le monde entier, et de grandir dans un large éventail d'habitats, y compris les environnements extrêmes tels que déserts ou les zones à forte concentration de sel ou de rayonnements ionisants, ainsi que dans sédiments marins profonds. Certains peuvent survivre à l'intense UV et rayonnement cosmique rencontré pendant Voyage espace. La plupart poussent dans des milieux terrestres, bien que plusieurs espèces vivent en partie ou uniquement dans les habitats aquatiques, comme le chytride Batrachochytrium dendrobatidis, un parasite qui a été responsable d'un déclin mondial les populations d'amphibiens. Cet organisme consacre une partie de son cycle de vie en tant que mobiles zoospore, lui permettant de se propulser dans l'eau et entrer son hôte amphibiens. D'autres exemples de champignons aquatiques comprennent ceux qui vivent dans zones hydrothermales de l'océan.
Environ 100 000 espèces de champignons ont été formellement décrite par taxonomistes, mais la biodiversité mondiale du royaume champignon ne est pas entièrement compris. Sur la base des observations du rapport entre le nombre d'espèces fongiques et le nombre d'espèces de plantes dans des milieux choisis, le règne fongique a été estimée à contenir environ 1,5 millions d'espèces; une récente (2011) estimation, il peut y avoir plus de 5 millions d'espèces. En mycologie, espèces ont toujours été distingués par une variété de méthodes et de concepts. Classification basée sur caractéristiques morphologiques, tels que la taille et la forme de spores ou des structures de fructification, a traditionnellement dominé taxonomie fongique. Les espèces peuvent également être distingués par leur biochimique et Caractéristiques physiologiques, tels que leur capacité à métaboliser certaines substances biochimiques, ou leur réaction à tests chimiques. Le concept d'espèce biologique discrimine les espèces en fonction de leur capacité à compagnon. L'application de outils moléculaires, tels que séquençage de l'ADN et l'analyse phylogénétique, pour étudier la diversité a grandement amélioré la résolution et ajouté robustesse des estimations de la diversité génétique au sein de divers groupes taxonomiques.
Morphologie
Structures microscopiques
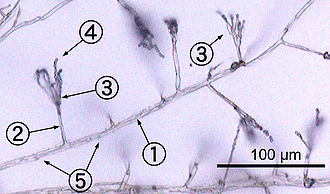
1. hyphe 2. 3 conidiophores. phialide 4. 5 conidies. cloisons
La plupart des champignons poussent comme hyphes, qui sont cylindriques, filiformes structures 2-10 um de diamètre et jusqu'à plusieurs centimètres de longueur. Hyphes se développent à leurs extrémités (apex); nouvelle hyphes sont généralement formé par émergence de nouvelles astuces long hyphes existante par un processus appelé ramification, ou parfois de plus en plus des conseils hyphes bifurquent (fourchette) donnant lieu à deux hyphes croissante parallèle. La combinaison de la croissance apicale et la ramification / bifurcation conduit à l'élaboration d'un mycélium, un réseau interconnecté de hyphes. Hyphes peut être soit cloisonnées ou coenocytic: hyphes cloisonnées sont divisés en compartiments séparés par des parois transversales (parois cellulaires internes, appelées cloisons, qui sont formés au perpendiculairement à la paroi de la cellule permet de donner sa forme hyphe), chaque compartiment contenant un ou plusieurs noyaux; hyphes coenocytic sont pas compartimenté. Cloisons ont pores qui permettent cytoplasme, organites, et parfois noyaux de passer à travers; un exemple est la cloison dolipore dans les champignons de l'embranchement Basidiomycota. Coenocytic hyphes sont essentiellement supercellules multinucléées.
De nombreuses espèces ont développé des structures de hyphes spécialisés pour l'absorption des nutriments à partir d'hôtes vivants; des exemples comprennent haustoria en espèces végétales-parasitaire de la plupart phylums fongiques, et arbuscules de plusieurs champignons mycorhiziens, qui pénètrent dans les cellules hôtes de consommer nutriments.
Bien que les champignons sont opisthokonts-un regroupement d'organismes connexes évolutif globalement caractérisée par une seule postérieure flagelle-tous les phylums l'exception de la chytrides ont perdu leurs flagelles postérieure. Les champignons sont inhabituel parmi les eucaryotes en ayant une paroi de cellule qui, en plus de glucanes (par exemple, β-1,3-glucane) et d'autres composants typiques, contient également la biopolymère de chitine.
Structures macroscopiques

Mycélium fongiques peuvent devenir visibles à l'œil nu, par exemple, sur différentes surfaces et des substrats, tels que des murs humides et sur la nourriture avariée, où ils sont communément appelés moules . Mycélium cultivés sur solide agar médias en laboratoire des boîtes de Pétri sont généralement désignés sous le nom de colonies. Ces colonies peuvent présenter des formes et des couleurs (en raison de spores ou la croissance de la pigmentation ) qui peuvent être utilisées en tant que fonctions de diagnostic pour l'identification des espèces ou groupes. Certaines colonies fongiques individuelles peuvent atteindre des dimensions et d'âges extraordinaires comme dans le cas d'un colonie clonale de Armillaria solidipes, qui se étend sur une superficie de plus de 900 ha (3,5 miles carrés), avec un âge estimé à près de 9000 années.
Le apothécie-une structure spécialisée important dans la reproduction sexuée dans les ascomycètes-est une forme de coupe- fructification qui détient la hyménium, une couche de tissu contenant les cellules sporifères. Les fructifications des basidiomycètes ( basidiocarpes) et certains ascomycètes peuvent parfois devenir très volumineux, et beaucoup sont bien connus comme des champignons .
La croissance et la physiologie

La croissance des champignons que hyphes sur ou dans des substrats solides ou sous forme de cellules uniques dans des environnements aquatiques est adapté pour l'extraction efficace des nutriments, parce que ces formes de croissance ont une grande aire de surface à des rapports en volume. Les hyphes sont spécifiquement conçus pour la croissance sur des surfaces solides, et d'envahir des substrats et des tissus. Ils peuvent exercer des efforts mécaniques importants pénétration; par exemple, la agent pathogène des plantes Magnaporthe grisea forme une structure appelée appressorium qui a évolué à la perforation des tissus végétaux. La pression générée par la appressorium, dirigée contre l'usine épiderme, ne peut dépasser huit mégapascals (1200 psi). Le champignon filamenteux Paecilomyces lilacinus utilise une structure similaire à pénétrer dans les oeufs de nématodes .
La pression mécanique exercée par la appressorium est généré à partir des processus physiologiques qui augmentent intracellulaire en produisant turgescence tels que les osmolytes glycérol. Adaptations morphologiques tels que ceux-ci sont complétés par enzymes hydrolytiques sécrétées dans l'environnement à digérer de grandes molécules organiques tels que- les polysaccharides, les protéines , les lipides et d'autres substrats organiques dans-molécules plus petites qui peuvent alors être absorbées en tant que nutriments. La grande majorité des champignons filamenteux grandir dans une polaire fashion-à-dire, par extension dans une direction par l'allongement à la pointe (apex) de l'hyphe. D'autres formes de la croissance fongique comprennent prolongement intercalaire (par exemple, par extension longitudinale des compartiments à hyphes qui sont au-dessous du sommet) comme dans le cas de certaines champignons endophytes, par croissance ou l'expansion de volume lors de l'élaboration de champignon stipes et autres grands organes. Croissance des champignons que structures multicellulaires consistant en somatiques et de la reproduction des cellules, une caractéristique évolué indépendamment chez les animaux et les plantes-a plusieurs fonctions, y compris le développement des fructifications pour la diffusion des spores sexuelles (voir ci-dessus) et biofilms substrat pour la colonisation et la communication intercellulaire.
Les champignons sont traditionnellement considérés hétérotrophes, les organismes qui se appuient uniquement sur carbone fixé par des organismes pour métabolisme. Les champignons ont évolué un degré élevé de polyvalence métabolique qui leur permet d'utiliser un large éventail de substrats organiques pour la croissance, y compris les composés simples tels que le nitrate , ammoniac , acétate, ou l'éthanol . Pour certaines espèces, il a été montré que le pigment mélanine peut jouer un rôle dans l'extraction de l'énergie à partir un rayonnement ionisant, tel que rayonnement gamma; Cependant, cette forme de La croissance "radiotrophic" a été décrite pour seulement quelques espèces, les effets sur les taux de croissance sont faibles, et le sous-jacent processus biophysiques et biochimiques ne sont pas connus. Les auteurs supposent que ce processus pourrait porter similitude Fixation du CO 2 via la lumière visible, mais en utilisant un rayonnement ionisant comme source d'énergie.
Reproduction

Reproduction fongique est complexe, reflétant les différences dans les modes de vie et génétique au sein de ce royaume diversifié d'organismes. On estime qu'un tiers de tous les champignons se reproduisent en utilisant plus d'une méthode de propagation; par exemple, la reproduction peut se produire en deux étapes bien différenciées au sein de la cycle de vie d'une espèce, la et la téléomorphe anamorphe. Conditions environnementales déclenchent génétiquement déterminés États développementistes qui conduisent à la création de structures spécialisées pour la reproduction sexuée ou asexuée. Ces reproduction de l'aide des structures de spores ou de spores contenant de dispersion efficace propagules.
La reproduction asexuée
La reproduction asexuée par spores végétatives ( conidies) ou par la fragmentation du mycélium est commun; elle maintient populations clonales adaptés à un particulier niche, et permet la dispersion plus rapide que la reproduction sexuée. Le "imperfecti de champignons" (champignons manque l'étape parfaite ou sexuel) ou Deuteromycota comprennent toutes les espèces qui ne ont pas un cycle sexuelle observable.
La reproduction sexuée
La reproduction sexuelle avec méiose existe dans tous les phylums fongiques (à l'exception de la Glomeromycota). Il diffère en de nombreux aspects de la reproduction sexuée chez les animaux ou les plantes. Des différences existent également entre les groupes fongiques et peuvent être utilisés pour différencier les espèces par des différences morphologiques dans les structures sexuelles et stratégies de reproduction. expériences d'accouplement entre les isolats fongiques peuvent identifier les espèces sur la base de concepts d'espèces biologiques. Les grands groupes fongiques ont d'abord été délimitées en fonction de la morphologie de leurs structures sexuelles et des spores; par exemple, les structures contenant des spores, asques et basides, peut être utilisé dans l'identification des Ascomycètes et les Basidiomycètes, respectivement. Certaines espèces peuvent permettre l'accouplement seulement entre les individus de face Type d'accouplement, tandis que d'autres peuvent se accoupler et se reproduire sexuellement avec une autre personne ou lui-même. Espèce de l'ex- système de reproduction sont appelés hétérothallique, et de celui-ci homothallique.
La plupart des champignons ont à la fois une haploïde et stade diploïde dans leurs cycles de vie. Dans reproduction sexuée des champignons, les personnes compatibles peuvent se combiner en fusionnant leurs hyphes ensemble dans un réseau interconnecté; ce processus, anastomose, est requise pour l'initiation du cycle sexuel. Ascomycètes et les Basidiomycètes passent par un dicaryotique stade, dans lequel les noyaux hérités des deux parents ne se combine pas immédiatement après la fusion cellulaire, mais demeurent distincts dans les cellules des hyphes (voir heterokaryosis).
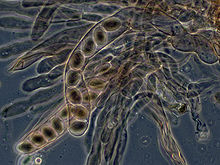
Dans ascomycètes, hyphes dicaryotique de la hyménium (la couche de tissu portant des spores) forment un crochet caractéristique au niveau du septum fongique. Pendant la division cellulaire, la formation du crochet assure une bonne répartition des noyaux nouvellement divisées dans les compartiments des hyphes apicales et basales. Un asque (ASCI pluriel) est alors formé, dans lequel caryogamie (fusion nucléaire) se produit. Les asques sont noyées dans un ascocarpe ou fructification. Caryogamie dans les asques est suivie immédiatement par la méiose et la production de ascospores. Après la dispersion, les ascospores peuvent germer et former un nouveau mycélium haploïde.
La reproduction sexuelle chez basidiomycètes est similaire à celle des ascomycètes. Compatible fusible hyphes haploïde pour produire un mycélium dicaryotique. Cependant, la phase dicaryotique est plus étendue dans les basidiomycètes, souvent également présente dans le mycélium en croissance végétative. Une structure anatomique spécialisé, appelé connexion de serrage, est formée à chaque cloison hyphes. Comme avec le crochet de structure similaire dans les ascomycètes, la liaison par serrage dans les basidiomycètes est nécessaire pour le transfert contrôlé de noyaux lors de la division cellulaire, pour maintenir la phase dicaryotique avec deux noyaux génétiquement différents dans chaque compartiment des hyphes. Un basidiocarpe est formée dans laquelle les structures du club-comme dit basides génèrent haploïde basidiospores après caryogamie et la méiose. Les basidiocarpes les plus connus sont les champignons, mais ils peuvent aussi prendre d'autres formes (voir Morphologie section).
Dans glomeromycetes (anciennement zygomycètes), hyphes haploïde de deux individus fusible, formant une Gamétange, une structure de cellule spécialisée qui devient fertile gamète cellule productrice. Le Gamétange se développe en une zygospore, une spore à paroi épaisse formée par l'union des gamètes. Lorsque le zygospore germe, il subit méiose, générer de nouvelles hyphes haploïde, qui peut ensuite former asexuée sporangiospores. Ces sporangiospores permettent au champignon de se disperser rapidement et germer dans de nouveaux mycélium fongiques haploïdes génétiquement identiques.
La dispersion des spores
Les deux spores ou sporangiospores asexués et sexués sont souvent dispersées activement par l'éjection de force de leurs structures reproductrices. Cette éjection assure sortie des spores des structures de reproduction ainsi que de voyager dans l'air sur de longues distances.

Spécialisées mécanismes mécaniques et physiologiques, ainsi que des structures de surface des spores (par exemple, hydrophobines), permettra spores efficace éjection. Par exemple, la structure de la spore portant des cellules de certaines espèces d'ascomycètes est telle que l'accumulation de substances affectant le volume des cellules et le bilan hydrique permet la décharge explosive de spores dans l'air. La décharge forcé de simples ballistospores spores appelées implique la formation d'une petite goutte d'eau (la chute de Buller), qui au contact de la spore conduit à sa sortie de projectile avec une accélération initiale de plus de 10 000 g; le résultat net est que la spore est éjecté de 0,01 à 0,02 cm, une distance suffisante pour qu'elle tombe par les branchies ou pores dans l'air ci-dessous. D'autres champignons, tels que la puffballs, se appuient sur des mécanismes alternatifs pour la libération des spores, telles que les forces mécaniques extérieures. Le Les champignons en nid d'oiseau utilisent la force de chute des gouttes d'eau pour libérer les spores de fructifications en forme de coupe. Une autre stratégie est vu dans le stinkhorns, un groupe de champignons avec des couleurs vives et odeur putride qui attirent les insectes pour disperser leurs spores.
Autres processus sexuels
Outre reproduction sexuelle régulière avec la méiose, certains champignons, tels que ceux dans les genres Et Penicillium Aspergillus, peuvent échanger du matériel génétique par l'intermédiaire parasexuel processus, initié par anastomose entre les hyphes et Plasmogamie de cellules fongiques. La fréquence et l'importance relative des événements parasexuel est pas claire et peut être inférieure à d'autres processus sexuels. Il est connu pour jouer un rôle dans l'hybridation intraspécifique et est susceptible nécessaire pour l'hybridation entre espèces, qui a été associée à des événements majeurs dans l'évolution fongique.
Évolution
Contrairement à plantes et animaux, le registre fossile début des champignons est maigre. Les facteurs qui contribuent probablement à la sous-représentation des espèces fongiques chez les fossiles comprennent la nature de fongique fructifications, qui sont doux, charnu, et facilement dégradables tissus et les dimensions microscopiques de la plupart des structures fongiques, qui sont donc pas tout à fait évident. Fossiles fongiques sont difficiles à distinguer de ceux d'autres microbes, et sont plus facilement identifiés quand ils ressemblent champignons existantes. Souvent récupéré d'une permineralized usine ou animal hôte, ces échantillons sont généralement étudiées par les préparatifs à section mince qui peuvent être examinées avec la microscopie optique ou La microscopie électronique à transmission. fossiles de compression sont étudiés en dissolvant la matrice environnante avec de l'acide, puis en utilisant la lumière ou microscopie électronique à balayage pour examiner les détails de surface.
Les premiers fossiles caractéristiques typiques de la date de champignons à l'possédant Eon Protérozoïque, certains 1430 il ya des millions d'années ( Ma); ces multicellulaire organismes benthiques avaient structures filamenteuses avec des cloisons, et étaient capables de l'anastomose. Des études plus récentes (2009) estiment l'arrivée d'organismes fongiques à environ 760 à 1060 Ma sur la base de comparaisons du taux de l'évolution dans les groupes étroitement liés. Pour une grande partie de la Ère paléozoïque (542 à 251 Ma), les champignons semblent avoir été aquatique et se composait des organismes similaires à l'existant chytrides en ayant spores du flagelle portant. L'adaptation évolutive à partir d'un milieu aquatique à un mode de vie terrestre a nécessité une diversification des stratégies écologiques pour l'obtention d'éléments nutritifs, y compris parasitisme, saprobism, et le développement de relations mutualistes tels que mycorhizes et lichénisation. (2009) Des études récentes suggèrent que l'état écologique ancestrale de la Ascomycota était saprobism, et que les indépendants lichénisation événements ont eu lieu à plusieurs reprises.
Les champignons probablement colonisé la terre pendant la Cambrian (542 à 488,3 Ma), bien avant que les plantes terrestres. Fossilisé hyphes et les spores récupéré de l' Ordovicien du Wisconsin (460 Ma) ressemblent moderne Glomerales et existaient à un moment où la flore terrestres susceptibles consistaient seulement non-vasculaire plantes de bryophytes comme. Prototaxites, qui était probablement un champignon ou de lichen, aurait été le plus grand organisme de la fin du Silurien . Fossiles fongiques ne deviennent pas commun et controverse jusqu'au début du Dévonien (416 à 359,2 Ma), quand ils sont abondants dans la Rhynie chert, principalement en Zygomycota et Chytridiomycota. Vers cette même époque, environ 400 Ma, le Ascomycota et Basidiomycota divergé, et tout le confort moderne classes de champignons étaient présents par la fin du Carbonifère ( Pennsylvanie, de 318,1 à 299 Ma).
Lichen les fossiles de ont été trouvés dans le Formation Doushantuo au sud de la Chine datant de 635 à 551 Ma. Les lichens sont une composante des écosystèmes terrestres début, et l'âge estimé de la plus ancienne lichen fossiles terrestre est de 400 Ma; Cette date correspond à l'âge de la plus ancienne connue sporocarpe fossiles, une espèce Paleopyrenomycites trouvés dans le Rhynie Chert. Le plus ancien fossile avec des caractéristiques microscopiques ressemblant basidiomycètes modernes est Palaeoancistrus, trouvé permineralized avec une fougère de la Pennsylvanie. Rare dans les archives fossiles sont les Homobasidiomycetes (un taxon à peu près équivalent aux espèces de champignons produisant de la Agaricomycetes). Deux ambre spécimens -preserved fournissent la preuve que les premiers champignons de champignons formant connu (les espèces disparues Archaeomarasmius leggetti) est apparu au cours de la mi- Crétacé , 90 Ma.
Quelque temps après la Événement Permien-Trias extinction (251,4 Ma), un pic fongique (à l'origine pensé pour être une abondance extraordinaire de spores fongiques dans les sédiments ) formé, ce qui suggère que les champignons étaient la forme de vie dominante à cette époque, ce qui représente près de 100% de la disposition des fossiles pour cette période. Toutefois, la proportion relative de spores fongiques par rapport aux spores formées par des algues espèces est difficile à évaluer, la pointe ne semble pas dans le monde entier, et dans de nombreux endroits, il ne est pas tombé sur la limite Permien-Trias.
Taxonomie
| Unikonta |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Bien que communément inclus dans la botanique programmes et manuels scolaires, les champignons sont plus étroitement liés à des animaux que de plantes et sont placés avec les animaux dans le groupe monophylétique de opisthokonts. Analyses aide phylogénie moléculaire soutiennent une origine monophylétique des Champignons. Le taxonomie des champignons est dans un état de flux constant, notamment en raison de la recherche récente basée sur des comparaisons d'ADN. Ces analyses phylogénétique actuelle renverser souvent classifications basées sur des méthodes plus anciennes et parfois moins discriminatoires sur la base de caractéristiques morphologiques et les concepts d'espèces biologiques obtenus à partir expérimentale accouplements.
Il n'y a pas système unique généralement reconnus aux niveaux taxonomiques élevés et il ya des changements de nom fréquents à tous les niveaux, à partir d'espèces vers le haut. Les efforts des chercheurs sont maintenant en cours pour établir et d'encourager l'utilisation d'un système unifié et plus cohérente nomenclature. Espèces fongiques peuvent également avoir des noms scientifiques multiples en fonction de leur cycle de vie et du mode (sexuée ou asexuée) de reproduction. Des sites Web tels que Index Fungorum et SITI Liste noms actuels des espèces fongiques (avec des renvois à synonymes âgées).
Le classement 2007 des champignons Uni est le résultat d'un effort de recherche collaborative à grande échelle impliquant des dizaines de mycologues et autres scientifiques travaillant sur la taxonomie fongique. Il reconnaît sept phylums, dont deux-Ascomycota et Basidiomycota-sont contenus dans une branche représentant sous-règne Dikarya. Le dessous cladogramme représente la majeure fongique taxons et leur relation à Opisthokonta et organismes Unikonta. Les longueurs des branches dans cet arbre ne sont pas proportionnelle à l'évolution distances.
Groupes taxonomiques
Le principal phylums (parfois appelés divisions) de champignons ont été classés principalement sur la base des caractéristiques de leur sexuelle structures reproductrices. Actuellement, sept phylums sont proposées: Microsporidia, Chytridiomycota, Blastocladiomycota, Neocallimastigomycota, Glomeromycota, Ascomycota et Basidiomycota.

L'analyse phylogénétique a démontré que le Microsporidia, parasites unicellulaires d'animaux et les protistes, sont assez récents et très dérivée champignons endobiotiques (vivant dans le tissu d'une autre espèce). Une étude réalisée en 2006 conclut que la Microsporidia sont un groupe sœur des champignons vrais; autrement dit, ils sont les plus proches de l'évolution par rapport à chaque autre.Hibbett et ses collègues suggèrent que cette analyse ne jure pas avec leur classement des Champignons, et bien que le Microsporidia sont élevés au statut d'embranchement, il est reconnu qu'une analyse plus poussée est nécessaire de clarifier les relations évolutives au sein de ce groupe.
Le Chytridiomycota sont communément connu comme chytrides. Ces champignons sont distribués dans le monde entier. Chytrides produisent zoospores qui sont capables de mouvement actif à travers les phases aqueuses avec un seul flagelle, conduisant début des taxonomistes de les classer comme protistes. la phylogénie moléculaire, présumées de séquences d'ARNr dans les ribosomes, suggèrent que les chytrides sont un groupe de base divergente de l'autre phylum fongique, comprenant de quatre grands clades avec des preuves suggestives pour paraphylie ou éventuellement polyphylie.
Le Blastocladiomycota ont été auparavant considéré comme un clade taxonomique dans le Chytridiomycota. Récentes données et moléculaires caractéristiques ultrastructurales, cependant, placent le blastocladiomycota comme un clade sœur de la Zygomycota, Glomeromycota et dikarya (Ascomycota et Basidiomycota). Les blastocladiomycetes sont saprotrophs, se nourrissant de matière organique en décomposition, et ils sont des parasites de tous les groupes eucaryotes. Contrairement à leurs proches parents, les chytrides, dont la plupart présentent la méiose zygotique, les blastocladiomycetes subissent la méiose sporic.
Le Neocallimastigomycota ont été précédemment placé dans le phylum Chytridomycota. Les membres de ce petit embranchement sont des organismes anaérobies, qui vivent dans le système digestif des grands mammifères herbivores et, éventuellement, dans d'autres environnements terrestres et aquatiques. Ils manquent de mitochondries , mais contiennent hydrogénosomes d'origine mitochondriale. Comme les chrytrids liés, forment neocallimastigomycetes zoospores qui sont uniflagellate ou postérieurement polyflagellate.
Les membres de la Forme Glomeromycota mycorhizes arbusculaires, une forme de symbiose dans laquelle hyphes envahissent les cellules des racines de la plante et les deux espèces bénéficient de l'augmentation de l'offre de nutriments résultant. Toutes les espèces connues Glomeromycota reproduisent de façon asexuée. L'association symbiotique entre le Glomeromycota et des plantes est ancienne, avec des preuves datant d'il ya 400 millions d'années. Anciennement partie de la Zygomycota (communément appelé «sucre» et «moules de pin»), l'Glomeromycota été élevé au statut de phylum en 2001 et remplace désormais le phylum âgé Zygomycota. Champignons qui ont été placés dans la Zygomycota sont maintenant réaffectés à la Glomeromycota, ou la sous-embranchements incertae sedis mucoromycotina, kickxellomycotina, le zoopagomycotina et la entomophthoromycotina. Quelques exemples bien connus de champignons autrefois dans la Zygomycota comprennent la moisissure noire de pain ( Rhizopus stolonifer ), et Pilobolus espèces, capable d'éjecter spores de plusieurs mètres dans les airs. Genres médicalement pertinents comprennent Mucor , Rhizomucor , et Rhizopus .
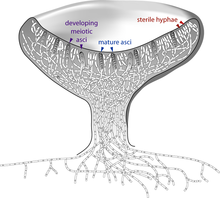
Le Ascomycota, communément connu comme les champignons de sac ou ascomycètes, constituent le plus grand groupe taxonomique dans le Eumycota. Ces champignons forment des spores méiotiques appelées ascospores, qui sont enfermés dans une structure en forme de sac spécial appelé asque. Ce phylum comprend morilles, quelques champignons et truffes, unicellulaires levures (par exemple, des genres Saccharomyces , Kluyveromyces , Pichia , et Candida ), et de nombreuses personnes vivant de champignons filamenteux comme saprotrophs, les parasites et symbiotes mutualistes. Genres éminents et importants de ascomycètes filamenteux comprennent Aspergillus , Penicillium , Fusarium et Claviceps . De nombreuses espèces d'ascomycètes ont seulement été observée subissant la reproduction asexuée (appelé espèces anamorphique), mais l'analyse des données moléculaires a souvent été en mesure d'identifier leurs proches dans le téléomorphes Ascomycota. Etant donné que les produits de la méiose sont retenus dans le asque en forme de sac, ascomycètes ont été utilisées pour élucider les principes de la génétique et l'hérédité (par exemple, Neurospora crassa ).
Les membres de la Basidiomycota, communément connu sous le nom du club ou les champignons basidiomycètes, produire méiospores appelés basidiospores sur des tiges de club, appelées basides. La plupart des champignons communs appartiennent à ce groupe, ainsi que la rouille et charbonneuses champignons, qui sont les principaux agents pathogènes des céréales. Autres basidiomycetes importants comprennent le maïs pathogène Ustilago maydis , les droits espèces commensales du genre Malassezia , et l' agent pathogène humain opportuniste, Cryptococcus neoformans .
organismes fongiformes
En raison de similitudes dans la morphologie et style de vie, les myxomycètes (les myxomycètes) et des moules d'eau (oomycètes) étaient auparavant classés dans le règne des Fungi. Contrairement aux champignons vrais les parois cellulaires de ces organismes contiennent de la cellulose et la chitine manquent. Myxomycetes sont unikonts comme des champignons, mais sont regroupés dans le Amoebozoa. Oomycètes sont diploïdes bikonts, regroupés dans le royaume Chromalveolate. Ni les moules d'eau ni myxomycètes sont étroitement liés aux véritables champignons, et, par conséquent, les taxonomistes groupe ne les champignons dans le royaume. Néanmoins, des études sur les oomycètes et myxomycètes sont encore souvent inclus dans les manuels de mycologie et de la littérature de recherche primaire.
Le Rozellida clade, y compris le "chytride" Rozella , est un groupe génétiquement disparate est surtout connu à partir de séquences d'ADN de l'environnement qui est un groupe de soeur aux champignons. Les membres du groupe qui ont été isolés manquent de la paroi cellulaire qui est caractéristique de chitine de champignons.
Le nucleariids, actuellement regroupés dans leChoanozoa, peuvent être le prochain groupe frère du clade de eumycete, et en tant que tels pourraient être inclus dans un royaume fongique élargi.
Écologie
Bien que souvent discrets, les champignons se produisent dans chaque environnement sur la Terre et jouent un rôle très important dans la plupart écosystèmes. Avec les bactéries, les champignons sont les principaux décomposeurs dans la plupart des écosystèmes terrestres (et certains aquatiques), et jouent donc un rôle essentiel dans les cycles biogéochimiques et dans de nombreux aliments nappes. Comme décomposeurs, ils jouent un rôle essentiel dans le cycle des nutriments, d'autant plus que saprotrophs et symbiotes, dégrader la matière organique des molécules inorganiques, qui peuvent ensuite réintégrer voies métaboliques anabolisants dans les plantes ou d'autres organismes.
Symbiose
Beaucoup de champignons ont d'importantesrelations symbiotiques avec les organismes de la plupart, sinon tous Royaumes.ces interactions peuvent êtremutualiste ou antagoniste dans la nature, ou dans le cas deschampignons commensaux sont d'aucun avantage apparent ou une atteinte à l'hôte.
Avec des plantes
Symbiose mycorhizienne entre les plantes et les champignons est l'une des associations les plantes-champignons les plus connus et est d'une importance significative pour la croissance des plantes et de la persistance dans de nombreux écosystèmes; plus de 90% de toutes les espèces végétales engager dans les relations avec les champignons mycorhiziens et sont tributaires de cette relation pour la survie.
La symbiose mycorhizienne est ancienne, datant de il ya au moins 400 millions d'années. Il augmente souvent l'absorption de l'usine de composés inorganiques, tels que le nitrate et le phosphate de sols ayant de faibles concentrations de ces éléments nutritifs essentiels. Les partenaires fongiques peuvent également servir de médiateur transfert de glucides et autres nutriments de plante à plante. Ces communautés mycorhiziens sont appelés «réseaux mycorhiziens communs". Un cas particulier de mycorhize est myco-hétérotrophie, où la plante parasite du champignon, l'obtention de l'ensemble de ses éléments nutritifs de son symbiote fongique. Certaines espèces fongiques habitent les tissus des racines à l'intérieur, les tiges, les feuilles et, dans ce cas, ils sont appelés endophytes. Similaire à mycorhizes, la colonisation par des champignons endophytes peut bénéficier deux symbiotes; par exemple, endophytes des graminées confèrent à leur hôte une résistance accrue aux herbivores et autres contraintes environnementales et reçoivent de la nourriture et un abri de la plante en retour.
Avec les algues et les cyanobactéries

Les lichens sont formés par une relation symbiotique entre les algues ou cyanobactéries (appelé dans la terminologie de lichen "photobiontes») et les champignons (surtout des diverses espèces d' ascomycètes et quelques basidiomycètes), dans laquelle les cellules photobionte individuelles sont intégrées dans un tissu formé par le champignon . Les lichens se produisent dans tous les écosystèmes sur tous les continents, jouent un rôle clé dans la formation du sol et l'initiation de la succession biologique et sont les formes de vie dominantes dans des environnements extrêmes, y compris polaire, alpine et les régions semi-arides du désert. Ils sont capables de se développer sur des surfaces défavorables, y compris les sols nus, les roches, l'écorce des arbres, bois, coquillages, balanes et les feuilles. Comme dans mycorhizes, la photobionte fournit des sucres et autres hydrates de carbone par la photosynthèse , tandis que le champignon fournit des minéraux et de l'eau. Les fonctions des deux organismes symbiotiques sont si étroitement liés qu'ils fonctionnent presque comme un organisme unique; dans la plupart des cas, l'organisme résultant est très différente de chacun des composants. Lichénisation est un mode commun de la nutrition; environ 20% des champignons entre 17.500 et 20.000 espèces décrites sont-lichénisés. Caractéristiques communes à la plupart des lichens comprennent l'obtention de carbone organique par photosynthèse, la croissance lente, de petite taille, longue durée de vie, de longue durée (de saison) structures de reproduction végétative, la nutrition minérale obtenus en grande partie de sources aéroportées, et une plus grande tolérance de la dessiccation que la plupart des autres organismes photosynthétiques dans le même habitat.
Avec insectes
Beaucoup d'insectes se livrent aussi à des relations de mutualisme avec les champignons. Plusieurs groupes de fourmis cultivent des champignons dans l'ordre Agaricales comme leur principale source de nourriture, alors que les scolytes cultivent diverses espèces de champignons dans l'écorce des arbres qu'ils infestent. De même, les femelles de plusieurs espèces de guêpes de bois (genre Sirex ) injectent leurs œufs avec des spores du champignon de bois pourrissant Amylostereum areolatum dans l' aubier du pin arbres; la croissance du champignon fournit des conditions nutritionnelles idéales pour le développement des larves de guêpe. termites sur le africaine savane sont également connus pour cultiver les champignons et levures des genres Candida et Lachancea habitent l' intestin d'un large éventail d'insectes, y compris neuroptères, coléoptères , et les cafards; on ne sait pas si ces champignons bénéficient leurs hôtes. Les larves de nombreuses familles de mouches fungicolous, en particulier celles de la superfamille Sciaroidea comme le Mycetophilidae et certains Keroplatidae nourrissent de fructifications fongiques et stérile mycorhizes.
Comme agents pathogènes et de parasites

Beaucoup de champignons sont des parasites sur les plantes, les animaux (y compris les humains), et d'autres champignons. Pathogènes graves de nombreuses plantes cultivées causant d'importants dégâts et pertes pour l'agriculture et la sylviculture sont les pyriculariose du riz Magnaporthe oryzae , agents pathogènes des arbres tels que Ophiostoma ulmi et Ophiostoma novo-ulmi qui causent la maladie hollandaise de l'orme, et Cryphonectria parasitica responsable de la brûlure du châtaignier, et agents pathogènes des plantes dans les genres Fusarium , Ustilago , Alternaria et Cochliobolus . Certains champignons carnivores, comme Paecilomyces lilacinus , sont les prédateurs des nématodes , qu'ils capturent en utilisant un tableau de structures spécialisées comme les anneaux de constriction ou filets adhésifs.
Certains champignons peuvent provoquer des maladies graves chez l'homme, dont plusieurs peuvent être fatales si non traitée. Il se agit notamment aspergilloses, candidoses, coccidioïdomycose, la cryptococcose, l'histoplasmose, mycétomes et paracoccidioïdomycose. En outre, les personnes immuno-déficiences sont particulièrement sensibles à la maladie par genres tels que Aspergillus , Candida , Cryptoccocus , Histoplasma , et la pneumonie . D'autres champignons peuvent attaquer les yeux, les ongles, les cheveux et la peau en particulier, les soi-disant champignons dermatophytes et kératinophiles, et causer des infections locales telles que la teigne et le pied d'athlète. Les spores fongiques sont également une cause d' allergies , et des champignons de différents groupes taxonomiques peuvent évoquer des réactions allergiques.
Usage humain
L'utilisation humaine des champignons pour la préparation ou la conservation des aliments et d'autres fins est vaste et a une longue histoire. Mushroom l'agriculture et de la cueillette des champignons sont les grandes industries dans de nombreux pays. L'étude des usages historiques et l'impact sociologique de champignons est connu comme ethnomycologie. En raison de la capacité de ce groupe à produire une vaste gamme de produits naturels avec antimicrobien ou d'autres activités biologiques, de nombreuses espèces ont longtemps été utilisés ou sont en cours de développement pour l'industrie de production d'antibiotiques, vitamines et anti-cancer et les médicaments anti-cholestérol. Plus récemment, des méthodes ont été développées pour le génie génétique des champignons, ce qui permet ingénierie métabolique des espèces fongiques. Par exemple, la modification génétique des espèces-levure qui sont faciles à cultiver à des taux rapides dans de grands navires-fermentation a ouvert les moyens de production pharmaceutique qui sont potentiellement plus efficace que la production par les organismes source d'origine.
Médicaments
De nombreuses espèces produisent des métabolites qui sont des sources importantes de pharmacologiquement médicaments actifs. Particulièrement importants sont les antibiotiques, y compris les pénicillines, un groupe de structure apparentée antibiotiques β-lactames qui sont synthétisés à partir de petits peptides. Bien que les pénicillines naturelles, telles que la pénicilline G (produit par Penicillium chrysogenum ) ont un spectre relativement étroit de l'activité biologique, un large éventail d'autres pénicillines peut être produit par modification chimique des pénicillines naturelles. Pénicillines modernes sont composés semi-synthétiques, obtenus initialement à partir de cultures de fermentation, mais structurellement modifiées pour des propriétés spécifiques souhaitables. D'autres antibiotiques produits par des champignons suivants: ciclosporine, couramment utilisé comme immunosuppresseur au cours de la chirurgie de transplantation; et l'acide fusidique, utilisé pour aider à l'infection par le contrôle de les bactéries résistantes à la méthicilline Staphylococcus aureus. l'utilisation généralisée de ces antibiotiques pour le traitement des maladies bactériennes, telles que la tuberculose , la syphilis, la lèpre , et beaucoup d'autres a commencé au début du 20ème siècle et continue de jouer un rôle majeur dans l'anti-bactérienne la chimiothérapie. Dans la nature, les antibiotiques de fongique ou bactérienne semblent jouer un double rôle: à des concentrations élevées, ils agissent comme défense chimique contre la concurrence avec d'autres micro-organismes dans riches en espèces environnements, tels que la rhizosphère, et à de faibles concentrations comme molécules quorum-sensing pour la signalisation intra ou interspécifique.
Autres médicaments produits par des champignons comprennent griséofulvine isolé de Penicillium griseofulvum , utilisé pour traiter les infections fongiques, et de statines ( inhibiteurs de la HMG-CoA réductase), utilisés pour inhiber la synthèse du cholestérol. Des exemples de statines trouvés dans les champignons comprennent mévastatine de Penicillium citrinum et de la lovastatine Aspergillus terreus et de la pleurote.
Aliments cultivés
La levure de boulanger ou Saccharomyces cerevisiae , un champignon unicellulaire, est utilisé pour faire du pain et d'autres produits à base de blé, comme la pâte à pizza et des boulettes. espèces de levure du genre Saccharomyces sont également utilisées pour produire des boissons alcoolisées par fermentation. Shoyu koji moule ( Aspergillus oryzae ) est un ingrédient essentiel dans le brassage de shoyu ( sauce de soja) et amour, et la préparation de miso, tandis que Rhizopus espèces sont utilisées pour la fabrication de tempeh. Plusieurs de ces champignons sont des espèces domestiquées qui ont été élevés ou sélectionnés en fonction de leur capacité à fermenter alimentaire sans produire mycotoxines nocives (voir ci-dessous), qui sont produites par de très étroitement liée Aspergilli . Quorn, un substitut de viande, est fabriqué à partir de Fusarium venenatum .
L'usage médical
 |  | |
Les champignons médicinaux Ganoderma lucidum(à gauche) et sinensis Ophiocordyceps(à droite) | ||
Certains champignons jouissent utilisation en tant qu'agents thérapeutiques dans des médicaments traditionnels, tels que la médecine traditionnelle chinoise. Champignons médicinaux notables avec une histoire bien documentée de l'utilisation comprennent subrufescens Agaricus , Ganoderma lucidum , et Ophiocordyceps sinensis . La recherche a identifié des composés produits par ces champignons et d'autres qui ont des effets biologiques inhibiteurs contre les virus et les cellules cancéreuses. Métabolites spécifiques, tels que le polysaccharide-K, l'ergotamine, et des antibiotiques β-lactamines, sont couramment utilisés en médecine clinique. Le shiitake est une source de lentinan, un médicament approuvé pour une utilisation clinique dans le traitement du cancer dans plusieurs pays, y compris le Japon . Dans l'Europe et le Japon , polysaccharide-K (nom de marque Krestin), un produit chimique dérivé de Trametes versicolor , est un approuvé adjuvant pour le traitement du cancer.
Espèces comestibles et toxiques

Les champignons comestibles sont des exemples bien connus de champignons. Beaucoup sont commercialement soulevé, mais d'autres doivent être récoltées à l'état sauvage. Agaricus bisporus , vendu comme champignons de Paris lorsque de petites ou portobello lorsque de plus grandes, est une espèce couramment consommés, utilisés dans des salades, des soupes, et de nombreux autres plats. Beaucoup de champignons asiatiques sont commercialement cultivées et ont gagné en popularité dans l'Ouest. Ils sont souvent disponibles frais dans les magasins et les marchés épiceries, y compris les champignons de paille ( Volvariella de volvacea ), pleurotes ( Pleurotus ostreatus ), shiitakes ( Lentinula edodes ), et enokitake ( Flammulina spp.).
Il ya beaucoup plus d'espèces de champignons qui sont prélevées dans la nature pour leur consommation personnelle ou de vente commerciale. champignons laitiers, morilles, chanterelles, truffes, trompettes de la mort, et porcini champignons ( Boletus edulis ) (aussi connu comme le bolet comestible) demande un prix élevé sur le marché. Ils sont souvent utilisés dans les plats gastronomiques.
Certains types de fromages nécessitent inoculation de lait caillé avec des espèces fongiques qui confèrent une saveur et une texture uniques pour le fromage. Des exemples comprennent le couleur bleue dans les fromages tels que le Stilton ou Roquefort, qui sont faites par inoculation de Penicillium roqueforti . Moules utilisés dans la production de fromage sont non-toxiques et sont donc sans danger pour la consommation humaine; Toutefois, les mycotoxines (par exemple, les aflatoxines, roquefortine C, patuline, ou autres) peuvent accumuler en raison de la croissance d'autres champignons durant la maturation ou le stockage du fromage.

De nombreuses espèces de champignons sont toxiques pour les humains, avec des toxicités allant de légers problèmes digestifs ou allergiques réactions ainsi que des hallucinations à des défaillances d'organes graves et la mort. Genres avec des champignons contenant des toxines mortelles comprennent Conocybe , Galerina , Lepiota , et le plus infâme, Amanita . Ce dernier genre comprend l'ange exterminateur ( A. virosa) et le bouchon de mort ( A. phalloïde) , la cause la plus fréquente d'intoxication aux champignons mortelle. Le faux morilles ( Gyromitra esculenta ) est parfois considéré comme une délicatesse lorsqu'il est cuit, mais peut être très toxiques lorsqu'ils sont consommés crus. Tricholoma equestre a été considéré comme comestible jusqu'à être impliqués dans des empoisonnements graves causant une rhabdomyolyse. Fly agaric ( Amanita muscaria ) aussi causer non occasionnelle intoxications mortelles, principalement à la suite de l'ingestion pour une utilisation comme drogue récréative pour ses propriétés hallucinogènes. Historiquement, amanite tue-mouche a été utilisé par les peuples différents en Europe et en Asie et de son utilisation actuelle pour religieuses ou fins chamaniques est rapporté par certains groupes ethniques tels que les personnes Koryak du nord-est Sibérie.
Comme il est difficile d'identifier avec précision un champignon sûr sans formation adéquate et des connaissances, il est souvent conseillé de supposer qu'un champignon sauvage est toxique et ne pas le consommer.
Antiparasitaire

Dans l'agriculture, les champignons peuvent être utiles si elles font activement concurrence pour les nutriments et l'espace avec des micro-organismes pathogènes tels que les bactéries ou d'autres champignons via le principe de l'exclusion compétitive, ou si elles sont des parasites de ces agents pathogènes. Par exemple, certaines espèces peuvent être utilisés pour éliminer ou supprimer la croissance des agents pathogènes des plantes nuisibles, tels que les insectes, les acariens, les mauvaises herbes , les nématodes et autres champignons qui provoquent des maladies de importantes plantes cultivées. Cela a généré un vif intérêt dans les applications pratiques qui utilisent ces champignons dans la lutte biologique contre ces ravageurs agricoles. champignons entomopathogènes peuvent être utilisés comme biopesticides, comme ils tuent activement insectes. Exemples qui ont été utilisés comme insecticides biologiques sont Beauveria bassiana, Metarhizium spp, Hirsutella spp, Paecilomyces ( Isaria ) spp, et Lecanicillium lecanii. les champignons endophytes des graminées du genre Neotyphodium , comme N. coenophialum , produire des alcaloïdes qui sont toxiques pour une gamme de vertébrés et d'invertébrés herbivores . Ces alcaloïdes protègent les plantes d'herbe de les herbivores , mais plusieurs alcaloïdes endophytes peuvent empoisonner les animaux de pâturage, comme les bovins et les ovins. Infecter les cultivars de pâturages ou les graminées fourragères avec Neotyphodium endophytes est une approche utilisée dans les programmes de sélection de l'herbe; les souches de champignons sont choisis pour produire seulement alcaloïdes qui augmentent la résistance aux insectes herbivores tels que, tout en étant non toxique pour le bétail.
Organismes modèles
Plusieurs découvertes pivots en biologie ont été faites par les chercheurs utilisant les champignons comme organismes modèles, qui est, les champignons qui poussent et sexuellement reproduisent rapidement dans le laboratoire. Par exemple, le un gène-une hypothèse de l'enzyme a été formulée par les scientifiques qui ont utilisé le moule à pain crassa Neurospora pour tester leur biochimique théories. Autres champignons modèles importants sont Aspergillus nidulans et les levures, Saccharomyces cerevisiae et Schizosaccharomyces pombe , dont chacun a une longue histoire d'utilisation pour étudier les questions en eucaryote biologie et de la cellule de la génétique , comme la régulation du cycle cellulaire, la structure de la chromatine, et la régulation des gènes. Autres modèles fongiques ont plus récemment émergé qui traitent chaque questions spécifiques biologiques pertinents à la médecine , pathologie des plantes, et les utilisations industrielles; des exemples comprennent Candida albicans , un pathogène dimorphisme opportuniste humaine, Magnaporthe grisea , un agent pathogène de la plante, et Pichia pastoris , une levure largement utilisé pour eucaryote expression de la protéine.
Autres
Les champignons sont largement utilisés pour produire des produits chimiques industriels tels que l'acide citrique, gluconique, lactique, les acides malique, et les enzymes industrielles, telles que les lipases utilisées dans les détergents biologiques, les cellulases utilisées dans la fabrication de cellulose et l'éthanol et des jeans délavés les amylases, les invertases, les protéases et les xylanases. Plusieurs espèces, et plus particulièrement les champignons psilocybine (familièrement connu comme les champignons magiques ), sont ingérés pour leurs propriétés psychédéliques, à la fois récréative et religieusement.
Les mycotoxines
![(6aR,9R)-N-((2R,5S,10aS,10bS)-5-benzyl-10b-hydroxy-2-methyl-3,6-dioxooctahydro-2H-oxazolo[3,2-a] pyrrolo[2,1-c]pyrazin-2-yl)-7-methyl-4,6,6a,7,8,9-hexahydroindolo[4,3-fg] quinoline-9-carboxamide](../../images/2778/277892.png)
Beaucoup de champignons produisent des composés biologiquement actifs, dont plusieurs sont toxiques pour les animaux ou les plantes et sont donc appelés mycotoxines. Un intérêt particulier pour les humains sont des mycotoxines produites par des moisissures qui causent la détérioration des aliments, et les champignons vénéneux (voir ci-dessus). Particulièrement infâme sont les mortels amatoxins dans certains Amanita les champignons, et les alcaloïdes de l'ergot de seigle, qui ont une longue histoire de causer de graves épidémies de l'ergotisme (le-feu de St Anthony) chez les personnes consommant de seigle ou liés céréales contaminées par des sclérotes de l'ergot de seigle, Claviceps purpurea . Autres mycotoxines notables incluent les aflatoxines, qui sont insidieuses toxines du foie et hautement métabolites cancérigènes produites par certains Aspergillus souvent des espèces qui poussent dans ou sur les céréales et les noix consommés par les humains, les ochratoxines, patuline, et trichothécènes (par exemple, Toxine T2) et les fumonisines, qui ont un impact significatif sur l'approvisionnement alimentaire humaine ou animale le bétail.
Les mycotoxines sont des métabolites secondaires (ou produits naturels), et la recherche a établi l'existence de voies biochimiques dans le seul but de produire des mycotoxines et autres produits naturels dans les champignons. Les mycotoxines peuvent fournir des avantages de conditionnement physique en termes d'adaptation physiologique, la concurrence avec d'autres microbes et les champignons, et la protection de la consommation ( fungivory).
Mycologie
Mycologie est la branche dela biologieconcernés par l'étude systématique des champignons, y compris leurs propriétés génétiques et biochimiques, leur taxonomie et leur utilisation pour les humains comme une source de médicaments, de nourriture etde substances psychotropes consommées à des fins religieuses, ainsi que leurs dangers , tels que l'empoisonnement ou d'une infection. Le domaine de la phytopathologie, l'étude des maladies des plantes, est étroitement liée parce que de nombreux agents pathogènes des plantes sont des champignons.

L'utilisation de champignons par l'homme remonte à la préhistoire; Ötzi, une momie bien conservé d'un 5300-year-old homme néolithique trouvé gelé dans les Alpes autrichiennes, effectué deux espèces de champignons polypore qui pourraient avoir été utilisés comme de l'amadou ( Amadouvier ), ou à des fins médicinales ( polypore du bouleau ). Les peuples anciens ont utilisé des champignons comme sources de nourriture, souvent sans le savoir-depuis des millénaires, dans la préparation du pain au levain et de jus fermentés. Certains des documents écrits les plus anciens contiennent des références à la destruction des cultures qui ont probablement été causée par des champignons pathogènes.
Histoire
Mycologie est une science relativement nouvelle qui est devenue systématique après le développement du microscope au 16e siècle. Bien que les spores fongiques ont d'abord été observés par Giambattista della Porta en 1588, l'ouvrage fondamental dans le développement de la mycologie est considéré comme la publication de 1729 travaux de Pier Antonio Micheli Nouvelle plantarum de genres . Micheli non seulement observer les spores, mais a montré que dans les conditions appropriées, ils pourraient être induits en plus en plus dans les mêmes espèces de champignons dont elles sont issues. Extension de l'utilisation du système binomial de la nomenclature introduite par Carl von Linné dans son plantarum espèces (1753), le Néerlandais Hendrik Christian Persoon (1761-1836) a créé la première classification des champignons avec une telle habileté afin d'être considéré comme l'un des fondateurs de la mycologie moderne . Plus tard, Fries Elias Magnus (1794-1878) a précisé la classification des champignons, en utilisant la couleur de spores et de diverses caractéristiques microscopiques, les méthodes utilisées par les taxonomistes encore aujourd'hui. Autres premiers contributeurs notables à la mycologie dans le 17ème à 19ème et début du 20ème siècles comprennent Miles Joseph Berkeley, Août Carl Joseph Corda, Anton de Bary, les frères Louis et René Charles Tulasne, Arthur RH Buller, Curtis G. Lloyd, et Pier Andrea Saccardo . Le 20ème siècle a vu une modernisation de la mycologie qui est venu des progrès de la biochimie , la génétique , la biologie moléculaire et la biotechnologie . L'utilisation de technologies de séquençage de l'ADN et l'analyse phylogénétique a fourni de nouvelles connaissances sur les relations fongiques et la biodiversité , et a contesté groupements basés sur la morphologie traditionnels dans fongique taxonomie.