

Sélection naturelle
Saviez-vous ...
SOS Enfants, qui se déroule près de 200 sos écoles dans le monde en développement, a organisé cette sélection. Visitez le site Web d'enfants SOS au http://www.soschildren.org/
La sélection naturelle est le processus par lequel favorable héréditaires traits deviennent plus fréquents dans successive générations d'une population de reproduction organismes , et les traits héréditaires défavorables devenus moins fréquents, en raison de la reproduction différentielle génotypes. La sélection naturelle agit sur le phénotype, ou les caractéristiques observables d'un organisme, tels que les personnes ayant des phénotypes favorables sont plus susceptibles de survivre et reproduire que celles des phénotypes moins favorables. Le phénotype génétique base, génotype associé avec le phénotype favorable, va augmenter dans fréquence sur les générations suivantes. Au fil du temps, ce processus peut entraîner adaptations qui se spécialisent notamment pour les organismes niches écologiques et peut éventuellement entraîner la apparition de nouvelles espèces. En d'autres termes, la sélection naturelle est le mécanisme par lequel l'évolution peut avoir lieu dans une population d'un organisme spécifique.
La sélection naturelle est une des pierres angulaires de moderne biologie . Le terme a été introduit par Charles Darwin dans son livre révolutionnaire 1859 L'origine des espèces dans laquelle la sélection naturelle a été décrit par analogie la sélection artificielle, un processus par lequel les animaux avec des traits jugés souhaitables par les éleveurs humains sont systématiquement privilégiés pour la reproduction. Le concept de la sélection naturelle a été développé à l'origine en l'absence d'une théorie valide de héritage; au moment de l'écriture de Darwin, on ne savait rien de modernes de la génétique . Bien que Gregor Mendel, le père de la génétique moderne, était un contemporain de Darwin, son travail se situerait dans l'obscurité jusqu'à ce que le début du 20e siècle. L'union de traditionnelle L'évolution darwinienne de découvertes ultérieures classique et génétique moléculaire est appelée la synthèse évolutionniste moderne. Bien que d'autres mécanismes de l'évolution moléculaire, tels que la théorie neutraliste avancé par Motoo Kimura, ont été identifiés comme des causes importantes de la diversité génétique, la sélection naturelle reste la principale explication unique pour l'évolution adaptative.
Principes généraux

La sélection naturelle agit sur l'organisme de phénotype, ou les caractéristiques physiques. Phénotype est déterminée par la constitution génétique d'un organisme ( génotype) et le milieu dans lequel vit l'organisme. Souvent, la sélection naturelle agit sur spécifique traits d'un individu, et les termes phénotype et du génotype sont utilisés strictement pour indiquer ces traits spécifiques.
Lorsque différents organismes dans une population possèdent différentes versions d'un gène pour un certain trait, chacune de ces versions est connu comme un allèle. Ce est cette variation génétique qui sous-tend traits phénotypiques. Un exemple typique est que certaines combinaisons de gènes de la couleur des yeux chez l'homme qui, par exemple, donnent lieu au phénotype des yeux bleus. (D'autre part, lorsque tous les organismes dans une part de la population le même allèle pour un caractère particulier, et cet état est stable dans le temps, l'allèle est dit être fixé dans cette population).
Certains traits sont régis par un seul gène, mais la plupart des caractères sont influencés par les interactions de nombreux gènes. Une variation dans l'un des nombreux gènes qui contribue à un trait peut avoir qu'un faible effet sur le phénotype; ensemble, ces gènes peuvent produire un continuum de valeurs phénotypiques possibles.
Nomenclature et l'utilisation
Le terme «sélection naturelle» a des définitions légèrement différentes dans des contextes différents. En termes simples, «sélection naturelle» est le plus souvent définie pour fonctionner sur les traits héréditaires, mais peut parfois se référer à la réussite de la reproduction différentielle des phénotypes indépendamment du fait que ces phénotypes sont héréditaires. La sélection naturelle est «aveugle» dans le sens que le niveau de succès de la reproduction des individus est une fonction du phénotype et non de savoir si ou dans quelle mesure ce phénotype est héréditaire. Après utilisation principale de Darwin le terme est souvent utilisé pour désigner à la fois la conséquence de la sélection aveugle et à ses mécanismes. Il est parfois utile de distinguer explicitement entre les mécanismes de sélection et ses effets; Lorsque cette distinction est importante, les scientifiques définissent «sélection naturelle» spécifiquement comme «les mécanismes qui contribuent à la sélection des individus qui se reproduisent», sans égard au fait que la base de la sélection est héréditaire. Ce est parfois dénommé «sélection naturelle phénotypique. '
Traits qui provoquent une plus grande réussite de la reproduction d'un organisme sont dits être sélectionné pour que ceux qui réduisent le succès sont sélectionnés contre. La sélection pour un trait peut également se traduire par la sélection d'autres traits corrélés qui ne se influencent pas directement remise en forme. Cela peut se produire à la suite de pléiotropie ou tringlerie de gène.
Aptitude
Le concept de remise en forme est au centre de la sélection naturelle. Cependant, comme avec la sélection naturelle ci-dessus, il ya d'importantes divergences d'opinion sur le sens précis du terme, et Richard Dawkins gère dans ses livres plus tard pour l'éviter complètement. (Il consacre un chapitre de son Phénotype étendu à discuter les différents sens dans lequel le terme est utilisé.) Bien que remise en forme est parfois familièrement comprise comme une qualité qui favorise la survie d'une personne en particulier - comme illustré dans la phrase bien connue survie du plus fort - la théorie moderne de l'évolution définit remise en forme en termes de reproduction individuelle. La base de cette approche est la suivante: si un organisme vit la moitié du temps que d'autres de son espèce, mais il a deux fois plus nombreux descendants survivant à l'âge adulte productive, ses gènes deviendra plus fréquent dans la population adulte de la prochaine génération. Ceci est connu comme reproduction différentielle.
Bien que la sélection naturelle agit sur les individus, son effet moyen sur tous les individus avec un génotype particulier correspond à l'aptitude de ce génotype. Très génotypes faible condition physique provoquer leurs porteurs d'avoir peu ou pas d'enfants en moyenne; Des exemples comprennent de nombreuses humain troubles génétiques comme la fibrose kystique . Conditions comme la drépanocytose peuvent avoir une faible remise en forme dans la population humaine en général, mais parce qu'il confère une immunité contre le paludisme, il a une valeur élevée de remise en forme dans les populations qui ont des taux d'infection élevés de paludisme. D'une manière générale, l'aptitude d'un organisme est une fonction des fitnesses de ses allèles. Depuis fitness est une quantité moyenne, cependant, il est possible une mutation favorable peut survenir chez un individu qui ne survit pas à l'âge adulte pour des raisons indépendantes.
Types de sélection
La sélection naturelle peut agir sur ne importe quel caractère phénotypique, et la pression sélective peut être produit par ne importe quel aspect de l'environnement, y compris compagnons et congénères, ou des membres de la même espèce. Toutefois, cela ne signifie pas que la sélection naturelle est toujours directionnelle et les résultats dans l'évolution adaptative; la sélection naturelle se traduit souvent par le maintien du statu quo en éliminant variantes moins bonne forme.
Le unité de sélection peut être l'individu ou il peut être un autre niveau de la hiérarchie d'organisation biologique, tels que les gènes, les cellules, et groupes de parenté. Il est toujours question de savoir si la sélection naturelle agit au niveau de des groupes ou des espèces afin de produire des adaptations qui profitent à un groupe plus large, non-parents. Sélection à un niveau différent tel que le gène peut conduire à une augmentation de l'aptitude à ce gène, tout en réduisant en même temps l'aptitude des individus porteurs de ce gène, dans un processus appelé intragénomique conflit. Dans l'ensemble, l'effet combiné de toutes les pressions de sélection à différents niveaux détermine la condition physique générale d'un individu, et donc le résultat de la sélection naturelle.
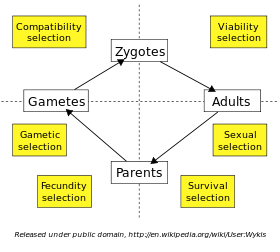
La sélection naturelle se produit à chaque étape de la vie d'un individu. Un organisme individuel doit survivre jusqu'à l'âge adulte avant de pouvoir reproduire, et la sélection de ceux qui atteignent ce stade est appelé sélection de viabilité. Chez de nombreuses espèces, les adultes doivent rivaliser avec l'autre pour les copains via la sélection sexuelle, et le succès dans cette compétition détermine qui sera la prochaine génération parent. Lorsque les individus peuvent se reproduire plus d'une fois, une survie plus longue dans la phase reproductive augmente le nombre de descendants, appelé sélection de survie. Le fécondité de deux femelles et les mâles (par exemple, le géant spermatozoïdes chez certaines espèces de Drosophila) peut être limitée par la sélection de la fécondité. La viabilité du produit gamètes peuvent différer, tout en intragénomique conflits tels que lecteur méiotique entre le gamètes haploïdes peuvent entraîner la sélection gamétique ou génique. Enfin, l'union de certaines combinaisons d'œufs et du sperme pourrait être plus compatibles que d'autres; On appelle cette sélection de compatibilité.
La sélection sexuelle
Il est également utile de faire une distinction entre mécaniste sélection écologique et le terme plus étroit la sélection sexuelle. de sélection écologique couvre tout mécanisme de la sélection en raison de l'environnement (y compris les parents, par exemple sélection de parentèle, et leurs congénères, par exemple la concurrence, infanticide), tandis que la sélection sexuelle se réfère spécifiquement à la concurrence entre congénères pour compagnons. La sélection sexuelle peut être intrasexuelle, comme dans les cas de concurrence entre individus du même sexe dans une population, ou intersexuelle, comme dans les cas où un sexe contrôle l'accès de reproduction en choisissant parmi une population de compagnons disponibles. Le plus souvent, la sélection intrasexuelle implique la concurrence mâle-mâle et la sélection intersexuelle implique le choix des femelles des mâles convenables, en raison de l'investissement généralement plus de ressources pour une femme que pour un homme en un seul organisme descendant. Cependant, certaines espèces présentent des rôles sexuels comportement inversé dans lequel ce sont les hommes qui sont les plus sélectifs dans le choix du partenaire; les exemples les plus connus de ce modèle se produisent dans certains poissons de la famille Syngnathidae, bien que les exemples probables ont également été trouvés dans amphibiens et oiseaux des espèces. Certaines fonctionnalités qui se limitent à un seul sexe d'une espèce particulière peuvent être expliqués par la sélection exercée par l'autre sexe dans le choix d'un partenaire, par exemple, le plumage extravagant de certains oiseaux mâles. De même, l'agression entre les membres du même sexe est parfois associée à des caractéristiques très distinctives, telles que les bois de cerfs, qui sont utilisés dans les combats avec d'autres cerfs. Plus généralement, la sélection intrasexuelle est souvent associée à dimorphisme sexuel, y compris les différences de taille entre les mâles et les femelles d'une espèce.
Un exemple: la résistance aux antibiotiques
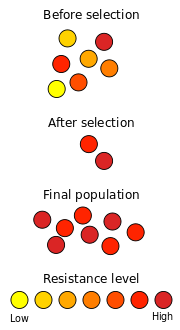
Un exemple bien connu de la sélection naturelle dans l'action est le développement de la résistance aux antibiotiques dans micro-organismes. Depuis la découverte de la pénicilline en 1928 par Alexander Fleming , des antibiotiques ont été utilisés pour lutter contre bactériennes maladies. Les populations naturelles de bactéries contiennent, entre leurs grands nombres de membres individuels, des variations considérables dans leur matériel génétique, principalement par suite de mutations. Lorsqu'elle est exposée aux antibiotiques, la plupart des bactéries meurent rapidement, mais certains peuvent avoir des mutations qui les rendent légèrement moins sensibles. Si l'exposition aux antibiotiques est courte, ces personnes survivent au traitement. Cette élimination sélective des individus inadaptés d'une population est la sélection naturelle.
Ces bactéries survivantes seront ensuite reproduire à nouveau, la production de la prochaine génération. En raison de l'élimination des individus mal adaptés à la dernière génération, cette population contient plus de bactéries qui ont une certaine résistance contre l'antibiotique. Dans le même temps, les nouvelles mutations se produisent, ce qui contribue nouvelle variation génétique à la variation génétique existant. Les mutations spontanées sont très rares, et les mutations avantageuses sont encore plus rares. Cependant, les populations de bactéries sont assez grand pour que quelques individus auront mutations bénéfiques. Si une nouvelle mutation réduit leur sensibilité à un antibiotique, ces personnes sont plus susceptibles de survivre à la prochaine fois confronté à cet antibiotique. Étant donné assez de temps, et une exposition répétée à l'antibiotique, une population de bactéries résistantes aux antibiotiques émergera.
L'utilisation généralisée et la mauvaise utilisation des antibiotiques a entraîné une augmentation de la résistance microbienne aux antibiotiques utilisés en clinique, au point que le Staphylococcus aureus résistant à la méticilline (SARM) a été décrit comme un ' superbactérie »en raison de la menace qu'il représente pour la santé et de son invulnérabilité par rapport aux médicaments existants. Les stratégies d'intervention comprennent généralement l'utilisation de différents antibiotiques plus puissants; Toutefois, nouvelle souches de SARM ont récemment émergé qui sont résistantes, même à ces médicaments. Ceci est un exemple de ce qui est connu comme un évolutif course aux armements, dans lequel les bactéries continuent à développer des souches qui sont moins sensibles aux antibiotiques, alors que les chercheurs médicaux continuent à développer de nouveaux antibiotiques qui peuvent les tuer. Une situation similaire se produit avec la résistance aux pesticides dans les plantes et les insectes. Les courses aux armements ne sont pas nécessairement induits par l'homme; un exemple bien documenté implique l'élaboration de la ARN voie d'interférence dans les plantes comme moyen de l'immunité innée contre les virus .
La théorie génétique de sélection naturelle
La sélection naturelle par lui-même est un concept simple, dans laquelle les différences entre les phénotypes de conditionnement physique jouent un rôle crucial. Ce est l'union de la sélection naturelle comme un mécanisme avec le matériel génétique comme un substrat qui offre la plupart des pouvoir explicatif de la théorie
Directionnalité de la sélection
Quand un composant d'un caractère est héréditaire, la sélection se modifie les fréquences de l'autre allèles ou des variantes du gène qui produit les variantes du trait. La sélection peut être divisée en trois classes, sur la base de son effet sur les fréquences des allèles.
Sélection directionnelle se produit quand un certain allèle a une plus grande aptitude que d'autres, ce qui entraîne une augmentation de la fréquence de cet allèle. Ce processus peut continuer jusqu'à l'allèle est fixe et l'ensemble de la population part le phénotype installateur. Il se agit d'une sélection directionnelle qui est illustré dans l'exemple de résistance aux antibiotiques ci-dessus .
Bien plus commune est stabilisation sélection (également connu sous le nom sélection de purification), ce qui réduit la fréquence des alleles qui ont un effet nuisible sur le phénotype - ce est-organismes produire de remise en forme inférieur. Ce processus peut continuer jusqu'à ce que l'allèle est éliminé de la population. Résultats de la sélection purification dans les caractéristiques génétiques fonctionnels, tels que les gènes codant pour des protéines ou séquences régulatrices, étant conservé au fil du temps en raison de la pression sélective contre les variants délétères.
Enfin, un certain nombre de formes de sélection équilibrante exister, qui ne entraîne pas de fixation, mais maintenir un allèle à des fréquences intermédiaires dans une population. Cela peut se produire dans espèces diploïdes (ce est ceux qui ont deux paires de chromosomes) lorsque individus hétérozygotes, qui ont différents allèles sur chaque chromosome en un seul locus génétique, avoir une condition physique supérieure à individus homozygotes qui ont deux des mêmes allèles. Cela se appelle avantage ou surdominance hétérozygote, dont l'exemple le plus connu est le paludisme résistance observée chez les humains hétérozygotes qui portent une seule copie du gène de la drépanocytose . Maintien de la variation allélique peut également se produire par sélection perturbateur ou la diversification, ce qui favorise les génotypes qui se écartent de la moyenne dans les deux sens (ce est le contraire de surdominance), et peut entraîner une distribution bimodale des valeurs de traits. Enfin, une sélection équilibrée peut se produire par sélection dépendant de la fréquence, où la remise en forme d'un phénotype particulier dépend de la distribution d'autres phénotypes dans la population. Les principes de la théorie des jeux ont été appliquées à comprendre les distributions de remise en forme dans ces situations, en particulier dans l'étude des sélection de parentèle et l'évolution de altruisme réciproque.
La sélection et la variation génétique
Une partie de toute la variation génétique est fonctionnellement neutre en ce qu'il ne produit aucun effet phénotypique ou de différence significative de remise en forme; l'hypothèse que représente cette variation pour une fraction importante de observée diversité génétique est connue comme la théorie neutraliste de l'évolution moléculaire et a été lancée par Motoo Kimura. Variation Neutre pensait autrefois pour englober plus de la variation génétique dans non codante de l'ADN, qui a été émis l'hypothèse d'être composée de " ADN poubelle ". Cependant, plus récemment, les rôles fonctionnels d'ADN non codant, tels que les fonctions de réglementation et de développement de ARN produits de gènes, a été étudiée en profondeur; une grande partie de la protéine non-codantes des séquences d'ADN sont hautement conservées sous sélection de purification forte et donc ne varient pas beaucoup d'un individu à, ce qui indique que des mutations dans ces régions ont des conséquences délétères. Lorsque la variation génétique ne entraîne pas de différences de remise en forme, la sélection ne peut pas affecter directement la fréquence d'une telle variation. En conséquence, la variation génétique sur ces sites sera plus élevé que dans les sites où la variation ne influence fitness.
l'équilibre de sélection de mutation
La sélection naturelle se traduit par la réduction de la variation génétique par l'élimination des individus inadaptés et par conséquent des mutations qui ont causé la mauvaise adaptation. Dans le même temps, les nouvelles mutations se produisent, ce qui entraîne une équilibre mutation-sélection. Le résultat exact de ces deux processus dépend à la fois sur la vitesse à laquelle de nouvelles mutations se produisent et de la force de la sélection naturelle, qui est une fonction de la façon défavorable la mutation se avère. Par conséquent, des changements dans le taux de mutation ou de la pression de sélection se traduira par une balance mutation-sélection différente.
Liaison génétique
Liaison génétique se produit lorsque le loci de deux allèles sont liés, ou à proximité immédiate de l'autre sur la chromosome. Lors de la formation de gamètes, recombinaison des résultats génétiques importantes dans redistribution des allèles. Cependant, il est probable qu'un tel remaniement se produit entre deux allèles dépend de la distance entre ces allèles; plus les allèles sont les uns aux autres, moins il est probable qu'un tel remaniement se produira. Par conséquent, lorsque la sélection un allèle cible, il en résulte automatiquement en sélection de l'autre allèle de même; grâce à ce mécanisme, la sélection peut avoir une forte influence sur les modèles de variation dans le génome.
Balayages sélectifs produisent quand un allèle devient plus fréquente dans une population en raison d'une sélection positive. Comme la prévalence de l'une des augmentations d'allèles, allèles liés peuvent également devenir plus commune, se ils sont neutres ou même légèrement délétère. Cela se appelle auto-stop génétique. A solides résultats de balayage sélectif dans une région du génome où le sélectionné de façon positive haplotype (l'allèle et ses voisins) sont essentiellement les seuls qui existent dans la population.
Que ce soit un balayage sélectif a eu lieu ou ne peut pas être étudiée par la mesure déséquilibre de liaison, ou si un haplotype donné est surreprésentée dans la population. Normalement, Résultats de la recombinaison génétique dans un remaniement des différents allèles dans un haplotype, et aucun des haplotypes vont dominer la population. Toutefois, au cours d'un balayage sélectif, la sélection d'un allèle spécifique se traduira également par sélection d'allèles voisins. Par conséquent, la présence de fort déséquilibre de liaison peut indiquer qu'il ya eu un balayage sélectif «récente», et cela peut être utilisé pour identifier les sites récemment sous sélection.
sélection de fond est le contraire d'un balayage sélectif. Si un site spécifique éprouve sélection purificatrice forte et persistante, la variation liée aura tendance à être éliminées avec elle, produisant une région dans le génome de la faible variabilité globale. Étant donné que la sélection d'arrière-plan est le résultat de nouvelles mutations délétères qui peuvent se produire de façon aléatoire dans tout haplotype, il ne produit pas de déséquilibre de liaison.
Evolution au moyen de la sélection naturelle
Une condition préalable à la sélection naturelle pour entraîner l'évolution adaptative, caractères nouveaux et spéciation, ce est la présence de héritable la variation génétique qui se traduit par des différences de fitness. La variation génétique est le résultat de mutations, recombinaisons et des altérations de la caryotype (le nombre, la forme, la taille et l'agencement interne de la chromosomes). Chacune de ces changements pourraient avoir un effet qui est très avantageuse ou très désavantageuse, mais des effets importants sont très rares. Dans le passé, la plupart des modifications dans le matériel génétique ont été considérés neutre ou proche de la neutralité parce qu'ils se sont produits dans ou ADN non codant conduit à une substitution synonyme. Toutefois, des recherches récentes suggèrent que de nombreuses mutations dans l'ADN non codant ne ont de légers effets délétères. Bien que les deux taux de mutation et les effets de mise en forme moyens des mutations dépendent de l'organisme, les estimations à partir de données à l'homme ont montré que la majorité des mutations sont légèrement délétère.

Par la définition de remise en forme, les personnes ayant plus de remise en forme sont plus susceptibles de contribuer descendants à la génération suivante, alors que les individus de moindre remise en forme sont plus susceptibles de mourir plus tôt ou ne parviennent pas à se reproduire. En conséquence, allèles sur le résultat moyen en plus grande forme physique deviennent plus abondants dans la prochaine génération, tout en allèles qui réduisent généralement remise en forme deviennent rares. Si les forces de sélection restent les mêmes depuis de nombreuses générations, allèles bénéfiques deviennent de plus en plus abondante, jusqu'à ce qu'ils dominent la population, tandis que les allèles avec une remise en forme moindre disparaissent. Dans chaque génération, de nouvelles mutations et recombinaisons apparaissent spontanément, produire un nouveau spectre de phénotypes. Par conséquent, chaque nouvelle génération sera enrichie par l'abondance croissante des allèles qui contribuent à ces traits qui ont été favorisés par la sélection, l'amélioration de ces traits plus de générations successives.
Certaines mutations se produisent dans ce qu'on appelle gènes régulateurs. Les variations de ceux-ci peuvent avoir des effets importants sur le phénotype de l'individu, car ils régulent la fonction de nombreux autres gènes. La plupart, mais pas tous, des mutations dans des gènes régulateurs entraînent non viable zygotes. Des exemples de mutations réglementaires non létales se produisent dans Les gènes Hox chez les humains, ce qui peut entraîner une côte cervicale ou polydactylie, une augmentation du nombre de doigts ou des orteils. Lorsque ces mutations entraînent une remise en forme plus élevée, la sélection naturelle favorisera ces phénotypes et le caractère nouveau ne se propage dans la population.
Traits établies ne sont pas immuables; traits qui ont une forte remise en forme dans un contexte environnemental peuvent être beaucoup moins en forme si les conditions environnementales changent. En l'absence de la sélection naturelle pour préserver un tel trait, il va devenir plus variable et se détériorer au fil du temps, qui pourrait causer un vestige manifestation du caractère. Dans de nombreux cas, la structure apparemment résiduelle peut conserver une fonctionnalité limitée, ou peut être co-opté pour d'autres traits avantageux dans un phénomène connu sous le nom préadaptation. Un exemple célèbre d'une structure résiduelle, l' oeil de la rat-taupe aveugle, est censé conserver fonction photopériode perception.
Spéciation
Spéciation nécessite l'accouplement sélective, qui se traduisent par une réduction flux de gènes. Accouplement sélectif peut être le résultat, par exemple, un changement dans l'environnement physique (isolement physique par une barrière extrinsèque), ou par sélection sexuelle entraîne homogamie. Au fil du temps, ces sous-groupes pourraient diverger radicalement pour devenir différentes espèces, soit en raison de différences de pressions de sélection sur les différents sous-groupes, soit parce que différentes mutations apparaissent spontanément dans les différentes populations, ou en raison de effets fondateurs - certains allèles potentiellement bénéfiques peuvent, par hasard, être présents dans seulement un ou l'autre des deux sous-groupes lors de leur première se séparent. Un mécanisme moins connu de la spéciation se produit par l'intermédiaire hybridation, bien documentée dans les plantes et parfois observée dans les groupes riches en espèces d'animaux tels que cichlidés. Ces mécanismes de spéciation rapide peuvent refléter un mécanisme de changement évolutif connu comme l'équilibre ponctué , ce qui suggère que le changement évolutif et en particulier la spéciation se produit généralement rapidement après l'interruption de longues périodes de stase.
Les changements génétiques au sein des groupes résultent dans l'augmentation incompatibilité entre les génomes des deux sous-groupes, réduisant ainsi le flux de gènes entre les groupes. Le flux de gènes cessera effectivement lorsque les mutations distinctifs caractérisant chaque sous-groupe se fixe. Aussi peu que deux mutations peuvent conduire à la spéciation: si chaque mutation a un effet neutre ou positif sur la condition physique quand ils se produisent séparément, mais un effet négatif quand ils se produisent ensemble, puis la fixation de ces gènes dans les sous-groupes respectifs mènera à deux isolement reproductif populations. Selon le concept de l'espèce biologique, celles-ci seront de deux espèces différentes.
Développement historique

Théories pré-darwiniennes
Plusieurs anciens philosophes ont exprimé l'idée que la nature produit une grande variété de créatures, apparemment au hasard, et que seules les créatures survivent que parviennent à subvenir à leurs besoins et de se reproduire avec succès; exemples bien connus comprennent Empédocle et son successeur intellectuelle, Lucrèce, tandis que des idées connexes ont ensuite été affinée par Aristote . La lutte pour l'existence a été décrit plus tard par al-Jahiz dans le 9ème siècle. Ces arguments classiques ont été réintroduits dans le 18ème siècle par Pierre Louis Maupertuis et d'autres, y compris Charles Darwin grand-père de Erasmus Darwin. Bien que ces précurseurs avaient une influence sur Darwinisme, ils avaient peu d'influence sur la suite de la trajectoire de la pensée évolutionniste de Charles Darwin.
Jusqu'au début du 19ème siècle, le vue dominant dans les sociétés occidentales est que les différences entre individus d'une espèce étaient inintéressants départs de leur Idéal platonicien (ou typus) de espèces créées. Cependant, la théorie de uniformitarianisme en géologie promu l'idée que les forces simples, faibles pourraient agir en continu sur de longues périodes de temps pour produire des changements radicaux dans le paysage de la Terre. Le succès de cette théorie a fait prendre conscience de la grande échelle de temps géologique et fait plausible l'idée que des changements minuscules, presque imperceptibles dans les générations successives pourraient avoir des conséquences sur l'échelle des différences entre les espèces. Début du 19e siècle évolutionnistes, tels que Jean Baptiste Lamarck suggéré hérédité des caractères acquis comme un mécanisme de changement évolutif; caractères adaptatifs acquis par un organisme au cours de sa durée de vie pourraient être héritées par la descendance de cet organisme, causant transformation des espèces. Cette théorie est venu à être connu sous le nom Lamarckisme et a eu une influence sur les idées anti-génétiques de la Stalinienne soviétique biologiste Trofim Lyssenko.
L'hypothèse de Darwin
Entre 1842 et 1844, Charles Darwin a exposé sa théorie de l'évolution par la sélection naturelle comme une explication pour l'adaptation et la spéciation. Il a défini la sélection naturelle comme le «principe selon lequel chaque légère variation [d'un trait], si ce est utile, est préservée". Le concept était simple mais puissant: individus les mieux adaptés à leurs environnements sont plus susceptibles de survivre et se reproduire. Tant qu'il y est une certaine variation entre eux, il y aura un choix inévitable de personnes avec les variations les plus avantageuses. Si les variations sont héritées, alors le succès reproductif différentiel conduira à une évolution progressive des populations particulières d'une espèce, et les populations qui évoluent être suffisamment différente pourrait éventuellement devenir des espèces différentes.
Les idées de Darwin ont été inspirés par les observations qu'il avait faites sur Le Voyage du Beagle, et par le travail de deux économistes. Le premier était Thomas Malthus , qui, dans Essai sur le principe de population, a noté que la population (si cochée) augmente de façon exponentielle alors que la production alimentaire croît seulement arithmétiquement; donc inévitables limites des ressources auraient des conséquences démographiques, conduisant à une «lutte pour l'existence», dans lequel seuls les plus forts survivront. La seconde était Adam Smith qui, La richesse des nations, a identifié un mécanisme de régulation des marchés libres, qu'il a appelé le " main invisible », ce qui suggère que les prix se ajuster selon l'offre et la demande. Ainsi, pour Darwin, la catastrophe qui devait se produire en fonction de Malthus a été tenu en échec et constamment améliorée par la concurrence (ou loi de la sélection).
Une fois la théorie a été formulée, Darwin était méticuleux au sujet de la collecte et de la preuve du raffinage, partageant ses idées seulement avec quelques amis; il a été inspiré à publier après le naturaliste Alfred Russel Wallace conçu indépendamment du principe et l'a décrit dans un essai qu'il a envoyé à Darwin. Un arrangement a été fait (à l'insu de Wallace) pour présenter son essai et deux courts écrits inédits de Darwin à la Linnean Society annonçant co-découverte du principe en Juillet 1858 Darwin a publié un compte rendu plus détaillé de son témoignage et les conclusions dans L'Origine des espèces en 1859. Dans la 6ème édition de L'origine des espèces de Darwin a reconnu que d'autres - notamment William Charles Wells en 1813, et Patrick Matthew en 1831 - avait proposé des théories similaires, mais ne les avait pas présenté entièrement ou dans des publications scientifiques notables.
Darwin pensait que la sélection naturelle par analogie à la façon dont les agriculteurs choisissent des cultures ou du bétail pour l'élevage, qu'il a appelé sélection artificielle; dans ses premiers manuscrits il se est référé à un «Nature» qui ferait la sélection. À l'époque, d'autres mécanismes de l'évolution, comme l'évolution par la dérive génétique ne étaient pas encore formulé explicitement, et Darwin réalisé que la sélection était probable qu'une partie de l'histoire: «Je suis convaincu qu '[il] a été les principaux, mais pas les moyens exclusifs de modification." Pour Darwin et ses contemporains, la sélection naturelle était donc essentiellement synonyme de l'évolution par la sélection naturelle . Après la publication de L'Origine des espèces, des gens instruits généralement admis que l'évolution se était produite dans une certaine forme. Cependant, la sélection naturelle est restée controversée en tant que mécanisme, en partie parce qu'il a été perçu comme trop faible pour expliquer l'éventail des caractéristiques observées des organismes vivants, et en partie parce que même les partisans de l'évolution rechigné à son «non guidé» et la nature non progressive, une response that has been characterized as the single most significant impediment to the idea's acceptance. Cependant, certains penseurs ont accueilli avec enthousiasme la sélection naturelle; après avoir lu Darwin, Herbert Spencer a introduit le terme survie du plus fort , qui est devenu un résumé populaire de la théorie. Bien que l'expression est encore souvent utilisé par des non-biologistes, les biologistes modernes éviter parce qu'elle est tautologique si plus fort est lu pour signifier fonctionnellement supérieure et est appliqué à des individus plutôt que considéré comme une quantité moyenne sur les populations. Dans une lettre à Charles Lyell en Septembre 1860, Darwin regrette l'utilisation du terme «sélection naturelle», préférant le terme «préservation de la nature».
Théorie synthétique de l'évolution
Seulement après l'intégration d'une théorie de l'évolution avec une appréciation statistique complexe du moine autrichien «redécouvertes» les lois de Gregor Mendel sur l' hérédité ne la sélection naturelle deviennent généralement acceptée par les scientifiques. Le travail de Ronald Fisher (qui a développé le langage des mathématiques et de la sélection naturelle en termes des processus génétiques sous-jacents), JBS Haldane (qui a introduit le concept de «coût» de la sélection naturelle), Sewall Wright (qui élucidé la nature de la sélection et l'adaptation), Theodosius Dobzhansky (qui a établi l'idée que la mutation, en créant génétique la diversité, fourni la matière première pour la sélection naturelle), William Hamilton (qui a conçu la sélection de parentèle), Ernst Mayr (qui a reconnu l'importance cruciale de l'isolement reproductif pour la spéciation) et beaucoup d'autres a formé la théorie synthétique de l'évolution. Cette synthèse a cimenté la sélection naturelle comme le fondement de la théorie de l'évolution, où il demeure aujourd'hui.
Impact de l'idée
Darwin les idées de ', avec ceux de Adam Smith et Karl Marx , ont eu une profonde influence sur la pensée du 19ème siècle. Peut-être la demande la plus radicale de la théorie de l'évolution par la sélection naturelle est que les «formes minutieusement construits, si différents les uns des autres, et dépendants les uns des autres d'une manière si complexe" évolué à partir de formes les plus simples de la vie de quelques principes simples. Cette affirmation a inspiré certains des plus ardents partisans de Darwin et a provoqué l'opposition la plus profonde. Le radicalisme de la sélection naturelle, selon Stephen Jay Gould , était en son pouvoir pour "détrôner certains des conforts les plus profondes et les plus traditionnels de la pensée occidentale". En particulier, il a contesté les croyances de longue date dans de tels concepts comme un endroit spécial et exalté pour l'homme dans le monde naturel et un créateur bienveillant dont les intentions ont été reflétées dans l'ordre et la conception de la nature.
La théorie sociale et psychologique
Les conséquences sociales de la théorie de l'évolution par la sélection naturelle sont également devenus la source de polémique continue. Friedrich Engels , un Allemand philosophe politique et co-auteur de l'idéologie du communisme , a écrit en 1872 que «Darwin ne savait pas ce une satire amère qu'il écrit sur l'humanité quand il a montré que la libre concurrence, la lutte pour l'existence, que les économistes célèbrent le succès historique le plus élevé, est l'état normal du règne animal ". Interprétation de la sélection naturelle comme nécessairement «progressiste», conduisant à augmenter 'avance' dans l'intelligence et de la civilisation, a été utilisé comme une justification de la colonisation et des politiques de l'eugénisme, ainsi que les positions socio-politiques plus larges maintenant décrit comme le darwinisme social. Konrad Lorenz a remporté le Nobel Prix de physiologie ou médecine en 1973 pour son analyse du comportement animal en termes du rôle de la sélection naturelle (en particulier de sélection de groupe). Cependant, en Allemagne, en 1940, dans les écrits qu'il a désavoués par la suite, il a utilisé la théorie comme une justification pour les politiques de l' nazie état. Il a écrit "... sélection pour la ténacité, l'héroïsme et l'utilité sociale ... qui doit être accompli par une institution humaine, si l'humanité, à défaut de facteurs sélectifs, ne doit pas être ruiné par la dégénérescence induite par la domestication. L'idée raciale la base de notre Etat a déjà accompli beaucoup de choses à cet égard. " D'autres ont développé des idées que les sociétés humaines et la culture évoluent par des mécanismes qui sont analogues à celles applicables à l'évolution des espèces.
Plus récemment, le travail parmi les anthropologues et les psychologues a conduit au développement de sociobiologie et plus tard la psychologie évolutionniste, un domaine qui tente d'expliquer les caractéristiques de la psychologie humaine en termes d'adaptation à l'environnement ancestral. L'exemple le plus frappant tels, notamment progressé dans les premiers travaux de Noam Chomsky et plus tard par Steven Pinker, est l'hypothèse que le cerveau humain est adapté pour acquérir les règles grammaticales de la langue naturelle. Autres aspects du comportement humain et des structures sociales, des normes culturelles spécifiques tels que l'inceste évitement pour des motifs plus larges tels que les rôles de genre, ont été émis l'hypothèse d'avoir des origines semblables adaptations à l'environnement au début de laquelle les humains modernes ont évolué. Par analogie à l'action de la sélection naturelle sur les gènes, le concept de mèmes - «unités de transmission culturelle", ou des équivalents de culture de gènes subissant la sélection et la recombinaison - a surgi, décrite pour la première sous cette forme par Richard Dawkins et ensuite élargi par les philosophes comme Daniel Dennett que des explications pour les activités culturelles complexes, y compris les droits conscience. des extensions de la théorie de la sélection naturelle à un tel éventail de phénomènes culturels ont été nettement controversée et ne sont pas largement acceptée.
Théorie de l'information et des systèmes
En 1922, Alfred Lotka a proposé que la sélection naturelle pourrait être comprise comme un principe physique qui pourrait être énergétiquement quantifié, un concept qui a été développé plus tard par Howard Odum que le principe de la puissance maximale par lesquelles les systèmes évolutifs avec avantage sélectif à maximiser le taux de transformation de l'énergie utile. Ces concepts sont parfois pertinentes dans l'étude des appliquées thermodynamique .
Les principes de la sélection naturelle ont inspiré une variété de techniques de calcul, telles que "douce" vie artificielle, qui simulent des processus sélectifs et peut être très efficace dans des entités «adaptation» à un environnement défini par une indication fonction de remise en forme. Par exemple, une classe d' heuristiques d'optimisation des algorithmes connus comme les algorithmes génétiques, mis au point par John Holland dans les années 1970 et élargi par David Goldberg, identifier les solutions optimales de reproduction simulée et la mutation d'une population de solutions définies par une première distribution de probabilité . Ces algorithmes sont particulièrement utiles lorsqu'il est appliqué à des problèmes dont la solution paysage est très rugueuse ou présente de nombreux minima locaux.