

Photosynthèse
À propos de ce écoles sélection Wikipedia
Enfants SOS offrent un chargement complet de la sélection pour les écoles pour une utilisation sur les intranets des écoles. Avant de vous décider à propos de parrainer un enfant, pourquoi ne pas en apprendre davantage sur différents organismes de parrainage premier ?
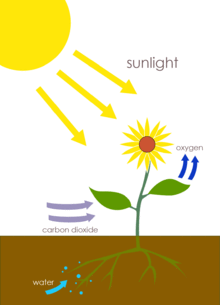
La photosynthèse est un processus par lequel la plupart des plantes et d'autres organismes captent l'énergie lumineuse du soleil pour conduire les processus chimiques nécessaires pour alimenter les organismes » métabolisme. Pour la plupart, ils combinent le dioxyde de carbone et l'eau pour former des hydrates de carbone , tels que des sucres. Cela donne lieu à la photosynthèse le nom, du grec [photographie], «lumière» et de σύνθεσις [synthèse], "mettre ensemble". Le processus libère également libres d'oxygène , la plupart du temps comme un produit des déchets. Cyanobactéries, la plupart des plantes , et la plupart des algues photosynthèse, et ces organismes sont appelés photoautotrophes. Photosynthèse maintient les niveaux d'oxygène atmosphérique et soit fournit l'énergie nécessaire pour la plupart la vie sur Terre, ou les précurseurs que les organismes peuvent métaboliser pour créer d'autres composés composés de carbone. Les principales exceptions sont chemotrophs, organismes qui gagnent de l'énergie par des réactions chimiques qui ne sont pas régis par l'énergie de la lumière.
Introduction
Bien que différentes espèces effectuer la photosynthèse de différentes manières, le processus commence toujours lorsque l'énergie de la lumière est absorbée par les structures de protéines molécules et des pigments. Ils sont appelés centres de réaction ; ils contiennent divers pigments, principalement vert chlorophylles. Dans les usines de ces protéines sont maintenus à l'intérieur organites appelés chloroplastes, principalement dans les cellules de la feuille. Pigments photosynthétiques dans les bactéries sont intégrés dans le membrane plasmique.
Dans ces réactions dépendant de la lumière une partie de l'énergie captée bandes électrons à partir de substances appropriées, généralement de l'eau, la séparation des atomes d'oxygène à partir de leurs ions et d'électrons d'hydrogène liés. Les atomes d'oxygène se combinent pour former des molécules d'oxygène et sont en grande partie éliminées, se échapper dans l'atmosphère. Les électrons et les ions hydrogène sont transférés à un composé appelé nicotinamide adénine dinucléotide phosphate (NADP +), réduisant à NADPH. La plupart du NADPH est consommée dans la production d'adénosine triphosphate (ATP), la «monnaie énergétique» des cellules.
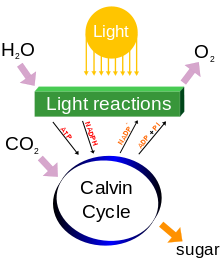
Chez les plantes, les algues et les cyanobactéries, les sucres sont produites à partir des produits de la photosynthèse immédiatement par une séquence de réactions luminescentes indépendant appelé le Cycle de Calvin. D'autres bactéries utilisent différents processus métaboliques, tels que la inverser cycle de Krebs. Dans le cycle de Calvin, le dioxyde de carbone dans l'atmosphère est incorporés dans des composés carbonés organiques déjà existants, tels que ribulose bisphosphate (RuBP). Cette incorporation initiale de carbone en composés organiques est appelé fixation du carbone. Utilisation de l'ATP et NADPH produite par les réactions dépendantes de la lumière, les composés résultants sont ensuite réduites en phosphate triose. Parmi tous les six molécules de triose phosphate produites, une est enlevée pour former d'autres hydrates de carbone et cinq sont «recyclée» dans le cycle de régénérer l'accepteur d'origine de dioxyde de carbone, RuBP.
Les premiers organismes photosynthétiques probablement évolué au début de la histoire de l'évolution de la vie. Les premiers organismes photosynthétiques probablement utilisés Des agents réducteurs tels que l'hydrogène ou sulfure d'hydrogène comme source d'électrons, plutôt que de l'eau. Les cyanobactéries oxydé que l'oxygène dans l'eau est apparue plus tard, l'excès d'oxygène et ils ont produit conduit à la catastrophe de l'oxygène, qui a finalement rendu la évolution de la vie complexe possible. Aujourd'hui, le taux moyen de capture d'énergie par la photosynthèse est globalement environ 130 térawatts, qui est environ six fois plus grand que le courant la consommation d'énergie de la civilisation. Dans le processus organismes photosynthétiques convertissent également autour de 100 à 115000 millions de tonnes (soit 100 à 115 pétagrammes) de carbone dans biomasse par an.
Vue d'ensemble
Les organismes photosynthétiques sont photoautotrophes, ce qui signifie qu'ils sont capables de synthétiser directement à partir de l'alimentation du dioxyde de carbone et de l'eau en utilisant l'énergie de la lumière. Cependant, tous les organismes qui utilisent la lumière comme source d'énergie portent pas la photosynthèse, puisque photohétérotrophes utilisent des composés organiques, plutôt que de dioxyde de carbone, en tant que source de carbone. Chez les plantes, les algues et les cyanobactéries, photosynthèse libère de l'oxygène. Cela se appelle la photosynthèse oxygénique. Bien qu'il existe des différences entre la photosynthèse oxygénique dans les plantes , les algues et cyanobactéries, l'ensemble du processus est assez similaire dans ces organismes. Cependant, il existe certains types de bactéries qui réalisent anoxygéniques photosynthèse, ce qui consomme du dioxyde de carbone, mais ne libère pas d'oxygène.
Le dioxyde de carbone est converti en sucres dans un processus appelé la fixation du carbone. La fixation du carbone est un endothermique réaction d'oxydoréduction, de sorte que la photosynthèse doit fournir à la fois une source d'énergie pour conduire ce processus, et les électrons nécessaire pour convertir le dioxyde de carbone dans un hydrate de carbone . Cet ajout d'électrons est un réaction de réduction. Dans les grandes lignes et en vigueur, la photosynthèse est le contraire de la respiration cellulaire, dans laquelle le glucose et d'autres composés sont oxydés pour produire du dioxyde de carbone et eau, et de libérer l'énergie chimique exothermique à conduire l'organisme de métabolisme. Cependant, les deux processus se déroulent à travers une séquence différente de réactions chimiques et dans différents compartiments cellulaires.
Le général l'équation pour la photosynthèse est donc:
- 2n + 2 CO 2 + 2n DH photons → 2 (CH 2 O) n 2n + DO
Le dioxyde de carbone + donneur d'électrons + énergie → lumière glucides + donneur d'électrons oxydé
Dans l'eau de la photosynthèse oxygénique est le donneur d'électrons et, depuis sa hydrolyse libère de l'oxygène, l'équation de ce processus est la suivante:
- 2n CO 2 + H 2 O 4n + photons → 2 (CH 2 O) n 2n + 2n O 2 + H 2 O
- dioxyde de carbone + eau + énergie lumineuse → glucides + oxygène + eau
Souvent, les molécules d'eau 2n sont annulés sur les deux côtés, ce qui donne:
- 2n CO 2 + H 2 O 2n + photons → 2 (CH 2 O) n 2n + O 2
- dioxyde de carbone + eau + énergie lumineuse → glucides + oxygène
D'autres procédés autres composés de substitution (par exemple, arsénite) pour l'eau dans le rôle d'alimentation en électrons; par exemple, certains microbes utilisent la lumière solaire pour oxyder l'arsénite de l'arséniate: L'équation de cette réaction est la suivante:
- CO 2 + (AsO 3- 3) + photons → (ASO 4 3-) + CO
- dioxyde de carbone + arsénite + énergie → arséniate + monoxyde de carbone lumière (utilisé pour construire d'autres composés dans les réactions ultérieures)
La photosynthèse a lieu en deux étapes. Dans la première étape, les réactions dépendant de la lumière ou des réactions légères captent l'énergie de la lumière et de l'utiliser pour faire les molécules de stockage d'énergie ATP et NADPH. Au cours de la seconde étape, les réactions de lumière indépendante utiliser ces produits pour capturer et réduire le dioxyde de carbone.
La plupart des organismes qui utilisent la photosynthèse pour produire l'utilisation d'oxygène la lumière visible de le faire, même si au moins trois utilisation ondes courtes infrarouge ou, plus précisément, le rayonnement rouge lointain.
Membranes et les organites photosynthétiques
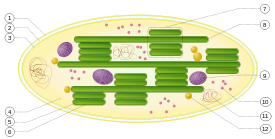
1. membrane externe
2. l'espace intermembranaire
3. membrane interne (1 + 2 + 3: enveloppe)
4. stroma (liquide aqueux)
5. thylacoïdiennes lumière (intérieur de thylakoïde)
6. membrane thylakoïde
7. granum (pile de thylakoïdes)
8. thylakoïde (lamelles)
9. l'amidon
10. ribosome
11. ADN plastidial
12. plastoglobule (chute de lipides)
Chez les bactéries photosynthétiques, les protéines qui prend la lumière pour la photosynthèse sont intégrés dans les membranes cellulaires, ce qui est la configuration la plus simple de ces protéines sont disposés. Toutefois, cette membrane peut être étroitement plié en feuilles cylindriques appelées thylakoïdes ou retroussé en rond vésicules appelées membranes intracytoplasmiques. Ces structures peuvent remplir plus de l'intérieur d'une cellule, la membrane donnant une très grande surface et donc d'augmenter la quantité de lumière que les bactéries peuvent absorber.
Chez les plantes et les algues, la photosynthèse a lieu dans organites appelés chloroplastes. Une caractéristique cellule végétale contient environ 10 à 100 chloroplastes. Le chloroplaste est entouré par une membrane. Cette membrane est constituée d'une membrane phospholipidique interne, une membrane externe de phospholipide, et un espace intermembranaire entre eux. Au sein de la membrane est un fluide aqueux appelé le stroma. Le stroma contient des piles (grana) de thylakoïdes, qui sont le site de la photosynthèse. Les thylakoïdes sont des disques aplatis, limitées par une membrane avec un lumen ou thylacoïdes espace intérieur. Le site de la photosynthèse est la membrane thylakoïde, qui contient intégrale et des complexes de protéines membranaires périphériques, y compris les pigments qui absorbent l'énergie lumineuse, qui forment les photosystèmes.
Les plantes absorbent la lumière en utilisant principalement le pigment chlorophylle, ce qui est la raison pour laquelle la plupart des plantes ont une couleur verte. Outre la chlorophylle, les plantes utilisent également des pigments tels que carotènes et xanthophylles. Les algues utilisent aussi de la chlorophylle, mais divers autres pigments sont présents que phycocyanine, carotènes, et xanthophylles dans algues vertes, phycoérythrine dans les algues rouges (rhodophytes) et fucoxanthine dans algues brunes et diatomées résultant en une grande variété de couleurs.
Ces pigments sont incorporés dans les plantes et les algues dans l'antenne-protéines spéciales. Dans ces protéines tous les pigments sont commandés à bien travailler ensemble. Une telle protéine est également appelée Antenne collectrice.
Bien que toutes les cellules dans les parties vertes de la plante ont chloroplastes, la plupart de l'énergie est capturée dans le laisse, sauf dans certaines espèces adaptées aux conditions de forte lumière du soleil et aridité, comme de nombreux Euphorbia et Cactus espèces, dont les principaux organes photosynthétique sont leurs tiges. Les cellules dans les tissus intérieurs d'une feuille, appelée mésophylle, peut contenir entre 450 000 et 800 000 chloroplastes pour chaque millimètre carré de feuille. La surface de la feuille est uniformément revêtu d'un résiste à l'eau, cireux cuticule qui protège la feuille de excessive évaporation de l'eau et diminue l'absorption de l'ultraviolet ou bleu lumière pour réduire le chauffage . Le transparent couche épiderme permet à la lumière de passer à travers la palissade cellules du mésophylle où la plupart de la photosynthèse a lieu.
Réactions légères
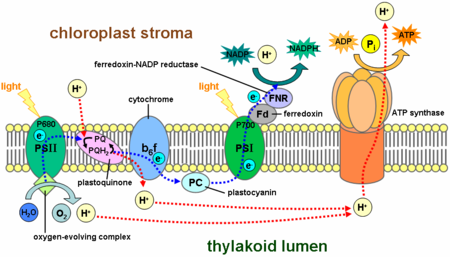
Dans le réactions claires, une molécule du pigment chlorophylle absorbe un photon et perd une électronique . Cet électron est passé à une forme modifiée de la chlorophylle appelé phéophytine, qui passe à l'électron à un molécule de quinone, ce qui permet le démarrage d'un flux d'électrons vers le bas un chaîne de transport d'électrons qui conduit à la réduction définitive de À NADP NADPH. En outre, cela crée un gradient de protons à travers la membrane du chloroplaste; sa dissipation est utilisé par ATP synthase pour la synthèse concomitante de l'ATP . La molécule de chlorophylle retrouve l'électron perdu d'une eau molécule par un processus appelé photolyse, qui libère un dioxygène (O 2) molécule. L'équation globale pour les réactions dépendant de la lumière dans les conditions de flux d'électrons non cyclique dans les plantes vertes est:
- 2 H 2 O + 2 + 3 + PNDA ADP + 3 P i + Lumière → 2 NADPH + H + 2 + 3 + O 2 ATP
Pas tout longueurs d'onde de la lumière peuvent soutenir la photosynthèse. Le spectre d'action photosynthétique dépend du type de pigments accessoires présents. Par exemple, dans les plantes vertes, la spectre d'action ressemble à la spectre d'absorption de chlorophylles et caroténoïdes avec des pics de bleu-violet et de la lumière rouge. Dans algue rouge, le spectre d'action se chevauche avec le spectre d'absorption phycobilines pour le bleu-vert lumière rouge, qui permet à ces algues de se développer dans les eaux plus profondes qui filtrent les longueurs d'onde utilisées par les plantes vertes. La partie non absorbée de la spectre de la lumière est ce qui donne leur couleur organismes photosynthétiques (par exemple, les plantes vertes, les algues rouges, bactéries pourpres) et est la moins efficace pour la photosynthèse dans les organismes respectifs.
Z régime
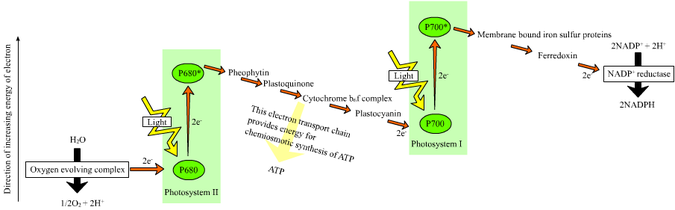
Chez les plantes, réactions dépendant de la lumière se produisent dans le membranes thylacoïdes de la chloroplastes et la consommation d'énergie de la lumière pour synthétiser l'ATP et NADPH. La réaction dépendant de la lumière a deux formes: cycliques et non-cycliques. Dans la réaction non cyclique, les photons sont capturés dans la lumière récolte les complexes d'antennes de photosystème II par chlorophylle et d'autres pigments accessoires (voir le schéma à droite). Quand une molécule de chlorophylle au coeur du centre photosystème II réactionnel obtenu énergie d'excitation suffisante des pigments d'antenne adjacents, un électron est transféré à la molécule accepteur d'électrons primaires, phéophytine, par un processus appelé séparation de charge photo-induit. Ces électrons sont ballottés par une chaîne de transport d'électrons, la soi-disant Z-schéma montré dans le diagramme, qu'initialement fonctions pour générer une chimiosmotique potentiel à travers la membrane. Une ATP synthase enzyme utilise le potentiel de faire chimiosmotique ATP au cours photophosphorylation, alors que NADPH est un produit de la borne réaction d'oxydoréduction dans le Z-régime. L'électron entre une molécule de chlorophylle dans Photosystème I. L'électron est excité en raison de la lumière absorbée par le photosystème. Un second porteur d'électrons accepte des électrons, qui est à nouveau transmis abaissement énergies de accepteurs d'électrons. L'énergie créée par les accepteurs d'électrons est utilisé pour déplacer des ions hydrogène à travers la membrane des thylakoïdes dans la lumière. L'électron est utilisé pour réduire la co-enzyme le NADP, qui a des fonctions à la réaction lumière indépendante. La réaction cyclique est similaire à celui de la non-cyclique, mais en diffère par la forme qu'elle génère uniquement ATP, de NADP et aucun réduite (NADPH) est créé. La réaction cyclique n'a lieu qu'à photosystème I. Une fois l'électron est déplacé de la photosystème, l'électron est passé les molécules accepteurs d'électrons et retourne à photosystème I, d'où il a été émis, d'où la réaction cyclique nom.
photolyse de l'eau
Le NADPH est le principal agent réducteur dans les chloroplastes, fournissant une source d'électrons énergétiques à d'autres réactions. Sa production chlorophylle laisse avec un déficit d'électrons (oxydé), qui doit être obtenu à partir d'un autre agent réducteur. Les électrons excités à partir de la perte en chlorophylle photosystème I est remplacé de la chaîne de transport d'électrons par plastocyanine. Toutefois, étant donné photosystème II comprend les premières étapes de la Z-régime, une source externe d'électrons est tenue de réduire ses molécules de chlorophylle a oxydés. La source d'électrons en vert-usine et la photosynthèse des cyanobactéries est l'eau. Deux molécules d'eau sont oxydés par quatre réactions de charges successives de séparation par photosystème II pour donner une molécule diatomique de l'oxygène et quatre hydrogène ions; le produit des électrons dans chaque étape est transféré à une activité rédox résidu de tyrosine qui réduit alors le jumelé chlorophylle photoxidized une espèce appelée P680 qui sert de primaire donneur d'électrons (à commande optique) dans le centre réactionnel du photosystème II. L'oxydation de l'eau est catalysée dans photosystème II par une structure ayant une activité rédox qui contient quatre manganèse ions et un ion calcium; cette complexe dégageant de l'oxygène se lie à deux molécules d'eau et stocke les quatre équivalents oxydants qui sont nécessaires pour mener la réaction d'oxydation dans l'eau. Photosystème II est le seul connu biologique enzyme qui effectue cette oxydation de l'eau. Les ions hydrogène contribuent au potentiel transmembranaire chimiosmotique qui conduit à la synthèse d'ATP. L'oxygène est un produit de déchets de réactions dépendant de la lumière, mais la majorité des organismes sur la Terre pour utiliser l'oxygène la respiration cellulaire, y compris les organismes photosynthétiques.
Réactions lumière indépendants
Cycle de Calvin
Dans le réactions légères indépendants (ou "sombres"), le enzyme RuBisCO capture de CO 2 de l' atmosphère et dans un processus qui nécessite du NADPH nouvellement formé, appelé le cycle de Calvin-Benson, libère sucres trois carbone, qui sont ensuite combinés pour former du saccharose et de l'amidon. L'équation globale pour les réactions légères indépendants dans les plantes vertes est:
- 3 CO 2 + 9 + 6 ATP + 6 NADPH + H → C 3 H 6 O 3-phosphate + ADP + 9 8 P i + 6 + NADP + 3 H 2 O
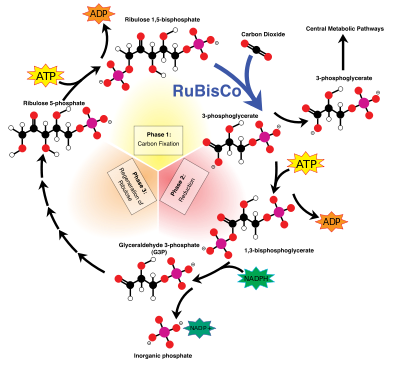
Pour être plus précis, la fixation du carbone produit un produit intermédiaire qui est ensuite converti en les produits finals de glucides. Les squelettes carbonés issus de la photosynthèse sont ensuite utilisés de diverses façons pour former d'autres composés organiques, tels que le matériau de construction la cellulose, en tant que précurseurs pour lipidique et l'acide aminé biosynthèse, ou comme combustible dans respiration cellulaire. Celui-ci se produit non seulement dans les plantes, mais également dans des animaux lorsque l'énergie à partir de plantes est transmis à travers un chaîne alimentaire.
La fixation ou la réduction du dioxyde de carbone est un procédé dans lequel du dioxyde de carbone se combine avec un sucre à cinq carbones, ribulose 1,5-bisphosphate (RuBP), pour donner deux molécules d'un composé à trois carbones, glycérate 3-phosphate (GP), également connu comme la 3-phosphoglycérate (PGA). GP, en présence d' ATP et NADPH à partir de la phase dépendant de la lumière, est réduite à la glycéraldéhyde 3-phosphate (G3P). Ce produit est également appelé 3-phosphoglycéraldéhyde ( PGAL) ou même phosphate comme triose. Triose est un sucre 3-carbone (voir glucides ). La plupart (5 sur 6 molécules) du G3P produite est utilisée pour régénérer RuBP de sorte que le processus peut se poursuivre (voir Cycle de Calvin-Benson). Les molécules 1 sur 6 des phosphates de triose pas «recyclés» condensent souvent pour former hexoses phosphates, qui donnent finalement le saccharose , amidon et cellulose. Les sucres produits au cours de carbone métabolisme squelettes rendement de carbone qui peuvent être utilisés pour d'autres réactions métaboliques comme la production d' acides aminés et des lipides .
concentrer les mécanismes de carbone
Sur terre
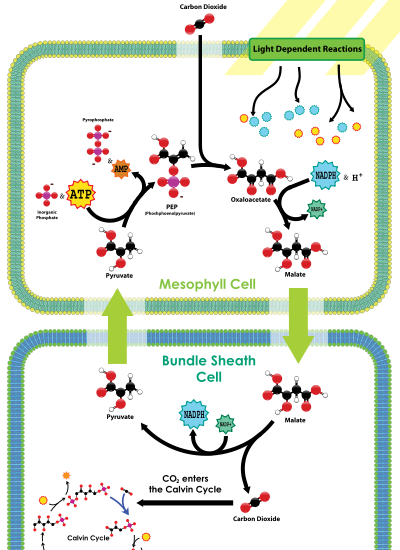
Dans des conditions chaudes et sèches, les plantes ferment leur stomates pour prévenir la perte d'eau. Dans ces conditions, CO 2 va diminuer, et de l'oxygène gazeux, produit par les réactions à la lumière de la photosynthèse, va diminuer dans la tige, pas les feuilles, provoquant une augmentation de par la photorespiration l'activité oxygenase de ribulose-1,5-bisphosphate carboxylase / oxygénase et la diminution de la fixation du carbone. Certaines plantes ont évolué mécanismes pour accroître la concentration de CO 2 dans les feuilles dans ces conditions.
C 4 plantes fixer chimiquement le dioxyde de carbone dans les cellules du mésophylle en l'ajoutant à la molécule à trois carbones phosphoénolpyruvate (PEP), une réaction catalysée par une enzyme appelée PEP carboxylase, la création de l'acide organique-quatre atomes de carbone l'acide oxaloacétique. Ou l'acide oxaloacétique malate synthétisée par ce processus est ensuite transporté vers spécialisée bundle cellules de la gaine où l'enzyme Enzymes du cycle Rubisco et autres Calvin se trouvent et où le CO 2 libérées par décarboxylation des acides quatre carbone est alors fixée par l'activité RuBisCO au sucre trois atomes de carbone Les acides 3-phosphoglycérique. La séparation physique de la RuBisCO de réactions lumineuses générant de l'oxygène réduit et augmente la photorespiration fixation du CO 2 et, par conséquent, la capacité photosynthétique de la feuille. C 4 plantes peuvent produire plus de sucre que C 3 plantes dans des conditions de forte lumière et la température. De nombreuses plantes cultivées importantes sont plantes C 4, notamment le maïs, le sorgho, la canne à sucre, et le millet. Les plantes qui ne utilisent pas PEP-carboxylase dans la fixation du carbone sont appelés C 3 plantes parce que la réaction de carboxylation primaire, catalysée par RuBisCO, produit le sucre à trois carbones acides 3-phosphoglycérique directement dans le cycle de Calvin-Benson. Plus de 90% des plantes C 3 utiliser la fixation du carbone, par rapport à 3% qui utilisent C 4 la fixation du carbone.
Xérophytes, comme les cactus et les plus succulentes, utilisent également la PEP carboxylase pour capturer le dioxyde de carbone dans un processus appelé Crassulacean métabolisme de l'acide (CAM). Contrairement à C 4 métabolisme, qui sépare physiquement la fixation du CO 2 à partir de PEP du cycle de Calvin, CAM sépare temporellement ces deux processus. Les plantes CAM ont une anatomie de C 3 plantes de feuille différent, et fixer le CO 2 la nuit, quand leurs stomates sont ouverts. Les plantes CAM stocker le CO 2 essentiellement sous la forme de l'acide malique par carboxylation de phosphoénolpyruvate en oxaloacétate, qui est ensuite réduit en malate. Décarboxylation du malate pendant la journée libère du CO 2 à l'intérieur des feuilles, permettant ainsi la fixation du carbone à 3-phosphoglycérate par RuBisCO. Seize mille espèces de plantes utilisent CAM.
Dans l'eau
Les cyanobactéries possèdent carboxysomes, qui augmentent la concentration de CO 2 autour de RuBisCO d'augmenter le taux de photosynthèse. Une enzyme, anhydrase carbonique, situé à l'intérieur de la carboxysome libère du CO 2 à partir des ions dissous hydrocarbonate (HCO 3 -). Avant le CO 2 diffuse hors il est rapidement épongé par RuBisCO, qui est concentrée dans les carboxysomes. HCO 3 - ions sont fabriqués à partir de CO 2 en dehors de la cellule par une autre et l'anhydrase carbonique sont activement pompé dans la cellule par une protéine de membrane. Ils ne peuvent pas traverser la membrane comme ils sont facturés, et dans le cytosol ils tournent le dos en CO 2 très lentement sans l'aide de l'anhydrase carbonique. Cela provoque le HCO 3 - ions à se accumuler dans la cellule d'où ils diffusent dans les carboxysomes. Pyrénoïdes dans les algues et anthocérotes agissent également se concentrer CO 2 autour rubisco.
Ordre et cinétique
L'ensemble du processus de la photosynthèse se déroule en quatre étapes:
| Étape | Description | Echelle de temps |
|---|---|---|
| 1 | Le transfert d'énergie antenne chlorophylle (membranes thylacoïdes) | femtosecondes pour picoseconde |
| 2 | Transfert d'électrons dans les réactions photochimiques (membranes thylacoïdes) | picoseconde nanoseconde |
| 3 | Electron chaîne de transport et la synthèse d'ATP (membranes thylacoïdes) | microseconde à milliseconde |
| 4 | La fixation du carbone et l'exportation de produits stables | milliseconde à deuxième |
Efficacité

Plantes convertissent la lumière en général l'énergie chimique d'un photosynthétique efficacité de 3-6%. L'efficacité photosynthétique des plantes réelles de la fréquence varie en fonction de la lumière étant converti, l'intensité lumineuse, la température et la proportion de dioxyde de carbone dans l'atmosphère, et peut varier de 0,1% à 8%. Par comparaison, panneaux solaires convertissent la lumière en l'énergie électrique avec un rendement d'environ 6-20% pour les panneaux fabriqués en série, et au-dessus de 40% dans des dispositifs de laboratoire.
systèmes de mesure de la photosynthèse ne sont pas conçus pour mesurer directement la quantité de lumière absorbée par la feuille. Néanmoins, les courbes de réponse à la lumière que les systèmes comme les produits LCpro-SD, ne permettent des comparaisons de l'efficacité photosynthétique entre les plantes.
Évolution

Les premiers systèmes de photosynthèse, tels que ceux de vert et pourpre et le soufre vert et bactéries pourpres non sulfureuses, sont considérés comme ayant été anoxygéniques, en utilisant diverses molécules comme Les donneurs d'électrons. Bactéries vert et violet soufre sont considérés comme ayant utilisé de l'hydrogène et du soufre comme un donneur d'électrons. Bactéries non sulfureuses vertes utilisé diverses aminés et autres des acides organiques. Bactéries pourpres non sulfureuses ont utilisé une variété de molécules organiques non spécifiques. L'utilisation de ces molécules est compatible avec la preuve géologique que l'atmosphère était très réduite à ce moment-là .
Les fossiles de ce que sont censées être organismes photosynthétiques filamenteuses ont été datés à l'âge de 3,4 milliards années.
La principale source d' oxygène dans l' atmosphère est photosynthèse oxygénique, et sa première apparition est parfois appelé le catastrophe de l'oxygène. Les données géologiques indiquent que la photosynthèse oxygénique, telle que celle en cyanobactéries, est devenu important au cours de la Ère paléoprotérozoïque il ya environ 2 milliards d'années. Photosynthèse moderne dans les plantes et les procaryotes photosynthétiques est plus oxygéné. Oxygenic photosynthèse utilise l'eau comme un donneur d'électrons, qui est oxydé à l'oxygène moléculaire (O 2) dans le centre réactionnel photosynthétique .
Symbiose et l'origine des chloroplastes
Plusieurs groupes d'animaux ont formé relations symbiotiques avec les algues photosynthétiques. Ce sont le plus commun dans coraux, éponges et anémones de mer. On suppose que cela est dû à la particulièrement simple plans d'organisation et de grandes surfaces de ces animaux par rapport à leur volume. En outre, quelques-uns marin mollusques Elysia viridis et Elysia chlorotica également à maintenir une relation symbiotique avec les chloroplastes qu'ils capturent des algues dans leur alimentation, puis stockent dans leur corps. Cela permet aux mollusques pour survivre uniquement par la photosynthèse pendant plusieurs mois à la fois. Certains des gènes de la plante noyau de la cellule ont même été transférés aux limaces, de sorte que les chloroplastes peuvent être fournis avec des protéines dont ils ont besoin pour survivre.
Une forme encore plus près de la symbiose peut expliquer l'origine des chloroplastes. Chloroplastes ont de nombreuses similitudes avec bactéries photosynthétiques, y compris une circulaire chromosome, de type procaryote ribosomes, et des protéines similaires dans le centre de réaction photosynthétique. Le théorie endosymbiotique suggère que les bactéries photosynthétiques ont été acquises (par endocytose) par les premiers eucaryotes cellules pour former les premières végétales cellules. Par conséquent, des chloroplastes peuvent être des bactéries photosynthétiques qui adaptés à la vie dans des cellules végétales. Comme les mitochondries , les chloroplastes possèdent encore leur propre ADN, distincte de la ADN nucléaire de leurs cellules hôtes de plante et les gènes de cet ADN chloroplastique ressemblent à celles cyanobactéries. ADN dans des chloroplastes de codes protéines redox tels que les centres de réaction photosynthétiques. Le CoRR hypothèse propose que cette coopération est nécessaire pour -location R edox R REGLEMENT.
Les cyanobactéries et de l'évolution de la photosynthèse
La capacité biochimique d'utiliser l'eau comme source d'électrons dans la photosynthèse a évolué une fois, dans un ancêtre commun existant cyanobactéries. Les données géologiques indiquent que cet événement de transformation a eu lieu tôt dans l'histoire de la Terre, il ya au moins 2450-2320 millions d'années (Ma), et il est supposé, beaucoup plus tôt. Les données disponibles à partir d'études de géobiologiques Archéennes (> 2500 Ma) roches sédimentaires indique que la vie existait 3500 Ma, mais la question de savoir quand la photosynthèse oxygénique évolué est toujours sans réponse. Une fenêtre paléontologique clair sur cyanobactéries évolution a ouvert environ 2000 Ma, révélant un biote déjà diversifié de bleu-verts. Les cyanobactéries est restée la première producteurs primaires à travers le Protérozoïque (2500-543 Ma), en partie parce que la structure de redox des océans favorisée photoautotrophes capables de fixation de l'azote . Les algues vertes rejoint bleu-verts comme les principaux producteurs primaires sur plateaux continentaux près de la fin de la Protérozoïque, mais seulement avec le Mésozoïque (251-65 Ma), radiations de dinoflagellés et les diatomées, coccolithophoridés fait la production primaire dans les eaux du plateau marines prendre forme moderne. Les cyanobactéries reste critique à les écosystèmes marins en tant que producteurs primaires dans tourbillons océaniques, comme des agents de la fixation biologique de l'azote, et, sous une forme modifiée, comme le plastes d'algues marines.
Une étude de 2010 par des chercheurs de Université de Tel Aviv a découvert que le Vespa orientalis (Vespa orientalis) convertit la lumière en énergie électrique en utilisant un pigment appelé xanthoptérine. Ce est la première preuve scientifique d'un membre du règne animal se engager dans la photosynthèse.
Découverte
Bien que certaines des étapes de la photosynthèse sont pas encore complètement compris, l'équation de la photosynthèse globale a été connu depuis le 19ème siècle.
Jan van Helmont a commencé la recherche du processus dans le milieu du 17e siècle quand il soigneusement mesuré la masse du sol utilisée par une usine et la masse de la plante comme il a grandi. Après avoir remarqué que la masse des sols très peu changé, il a émis l'hypothèse que la masse de la plante en croissance doit venir de l'eau, la seule substance at-il ajouté à la plante en pot. Son hypothèse était exacte partiellement - une grande partie de la masse acquise vient aussi du dioxyde de carbone ainsi que de l'eau. Cependant, ce était un point à l'idée de signalisation que la majeure partie d'une plante est biomasse provient des entrées de la photosynthèse, et non pas le sol lui-même.
Joseph Priestley, un chimiste et le ministre, a découvert que, quand il a isolé un volume d'air sous un pot inversé, et brûlé une bougie en elle, la bougie se brûler très vite, bien avant qu'il a manqué de cire. Il a en outre découvert que une souris pourrait de même "blesser" l'air. Il a ensuite montré que l'air qui avait été «lésé» par la bougie et la souris pourrait être rétablie par une plante.
En 1778, Jan Ingenhousz, médecin de la cour à l' autrichienne impératrice, a répété les expériences de Priestley. Il a découvert que ce était l'influence de la lumière du soleil sur la plante qui pourrait l'amener à revivre une souris dans une question d'heures.
En 1796, Jean Senebier, un Suisse pasteur, botaniste et naturaliste, a démontré que les plantes vertes consomment du dioxyde de carbone et rejettent de l'oxygène sous l'influence de la lumière. Peu de temps après, Nicolas-Théodore de Saussure a montré que l'augmentation de la masse de la plante pendant sa croissance pourrait ne pas être uniquement due à l'absorption de CO 2 mais aussi à l'incorporation de l'eau. Ainsi, la réaction de base par photosynthèse qui est utilisé pour produire des denrées alimentaires (tels que le glucose) a été décrite.
Cornelis Van Niel fait des découvertes clés expliquant la chimie de la photosynthèse. En étudiant bactéries de soufre pourpres et bactéries vertes il était le premier scientifique à démontrer que la photosynthèse est un dépendant de la lumière réaction d'oxydoréduction, dans lequel de l'hydrogène réduit le dioxyde de carbone.
Robert Emerson découvert deux réactions à la lumière de la productivité de l'usine de test en utilisant différentes longueurs d'onde de la lumière. Avec le rouge seul, les réactions à la lumière ont été supprimées. Si le fil bleu et rouge ont été combinées, la sortie était beaucoup plus importante. Ainsi, il y avait deux photosystèmes, une absorber jusqu'à 600 longueurs d'onde nm, l'autre à 700 nm. Le premier est connu comme PSII, ce dernier est PSI. PSI ne contient que la chlorophylle a, PSII contient principalement chlorophylle a avec la plupart de la chlorophylle b disponibles, entre autre pigment. Il se agit notamment phycobilines, qui sont les pigments rouges et bleus d'algues rouges et bleu respectivement, et fucoxanthol pour les algues brunes et les diatomées. Le processus est plus productive lorsque l'absorption de quanta sont égaux à la fois dans le PSII et PSI, assurant que l'énergie d'entrée du complexe de l'antenne est divisé entre le PSI et le système PSII, qui à son tour la photochimie.

Robert Hill pense qu'un complexe de réactions consistant en un intermédiaire de cytochrome b 6 (maintenant un plastoquinone), une autre est de cytochrome f à une étape dans les mécanismes de génération d'hydrates de carbone. Ils sont liés par la plastoquinone, qui ne nécessite de l'énergie pour réduire le cytochrome f car ce est un agent réducteur suffisante. D'autres expériences pour prouver que l'oxygène développé au cours de la photosynthèse des plantes vertes est venu de l'eau, ont été réalisées par Hill en 1937 et 1939. Il a montré que isolée chloroplastes dégagent de l'oxygène en présence d'agents réducteurs non naturels tels que le fer oxalate, ferricyanure ou benzoquinone après l'exposition à la lumière. La réaction de Hill est la suivante:
- 2 H 2 O + 2 + A (lumière, les chloroplastes) → 2 H 2 + O 2
où A est l'accepteur d'électrons. Par conséquent, à la lumière, l'accepteur d'électrons est réduite et l'oxygène se dégage.
Samuel Ruben et Martin Kamen utilisé isotopes radioactifs pour déterminer que l'oxygène libéré dans la photosynthèse est venu de l'eau.
Melvin Calvin et Andrew Benson, avec James Bassham, élucidé la voie de l'assimilation du carbone (le cycle photosynthétique de réduction de carbone) dans les plantes. Le cycle de réduction de carbone est connue sous le nom Cycle de Calvin, qui ignore la contribution de Bassham et Benson. De nombreux scientifiques se réfèrent au cycle le cycle de Calvin-Benson, Benson-Calvin, et certains ont même l'appellent le cycle de Calvin-Benson-Bassham (ou CBB).
Scientifique lauréat du prix Nobel Rudolph Marcus a pu découvrir la fonction et la signification de la chaîne de transport d'électrons.
Otto Heinrich Warburg et Dean Burk a découvert la réaction de photosynthèse I-quantique qui sépare le CO2, activé par la respiration.
Louis NM Duysens etJan Amesz découvert que la chlorophylle a va absorber une lumière, oxyder cytochrome f, la chlorophylle a (et d'autres pigments) va absorber une autre lumière, mais permettra de réduire ce même cytochrome oxydée, indiquant les deux réactions légères sont en série.
Facteurs

Il ya trois principaux facteurs affectant la photosynthèse et plusieurs facteurs corollaires. Les trois principaux sont:
- Lumière irradiance et longueur d'ondes
- Le dioxyde de carbone concentration
- Température.
Intensité de lumière (irradiance), longueur d'onde et de la température
Au début du 20e siècle, Frederick Blackman etGabrielle Matthaei étudié les effets de l'intensité lumineuse (irradiance) et de la température sur le taux d'assimilation de carbone.
- A température constante, le taux d'assimilation de carbone varie en fonction de l'irradiance, augmentant d'abord que les éclairement augmente. Cependant, à irradiance élevée, cette relation ne tient plus et le taux d'assimilation de carbone atteint un plateau.
- Au rayonnement constante, le taux d'assimilation de carbone augmente à mesure que la température augmente dans une gamme limitée. Cet effet est observé seulement à des niveaux d'éclairement élevés. A faible rayonnement, augmentant la température a peu d'influence sur le taux d'assimilation de carbone.
Ces deux expériences illustrent des points essentiels: d'abord, à partir de la recherche, il est connu que, en général, des réactions photochimiques sont pas affectés par la température . Cependant, ces expériences montrent clairement que la température affecte le taux d'assimilation de carbone, il doit y avoir deux séries de réactions dans le processus complet de l'assimilation du carbone. Ce sont, bien sûr, le stade dépendant de la lumière 'photochimique »et le stade de lumière indépendante, dépendant de la température. Deuxièmement, les expériences de Blackman illustrent le concept de facteurs limitants. Un autre facteur limitant est la longueur d'onde de la lumière. Les cyanobactéries, qui réside à plusieurs mètres sous l'eau, ne peut pas recevoir les longueurs d'onde correctes nécessaires pour provoquer la séparation de charge photoinduite dans les pigments photosynthétiques classiques. Pour lutter contre ce problème, une série de protéines avec différents pigments entourent le centre de réaction. Cette unité est appelée phycobilisome.
les niveaux de dioxyde de carbone et photorespiration
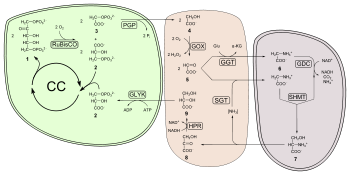
Comme les concentrations de dioxyde de carbone augmente, la vitesse à laquelle les sucres sont faites par les réactions de lumière indépendante augmente jusqu'à limitée par d'autres facteurs. RuBisCO, l'enzyme qui capture du dioxyde de carbone dans les réactions de lumière indépendante, a une affinité de liaison pour à la fois le dioxyde de carbone et l'oxygène. Lorsque la concentration de dioxyde de carbone est élevé, RuBisCO se fixer le dioxyde de carbone. Cependant, si la concentration en dioxyde de carbone est faible, la RuBisCO se lier à la place de l'oxygène du dioxyde de carbone. Ce processus, appelé photorespiration, utilise de l'énergie, mais ne produit pas de sucres.
Activité oxygénase RuBisCO est désavantageux pour les plantes, pour plusieurs raisons:
- Un produit de l'activité de l'oxygénase est phosphoglycolate (2 carbone) au lieu du 3-phosphoglycérate (3 carbone). Phosphoglycolate ne peut pas être métabolisé par le cycle de Calvin-Benson et représente la perte de carbone du cycle. Une forte activité oxygénase, donc, draine les sucres qui sont nécessaires pour recycler ribulose 5-bisphosphate et pour la poursuite du cycle de Calvin-Benson.
- Phosphoglycolate est rapidement métabolisé en glycolate qui est toxique pour une plante à une concentration élevée; il inhibe la photosynthèse.
- La récupération du glycolate est un processus coûteux énergétiquement qui utilise la voie de glycolate, et seulement 75% du carbone est renvoyée au cycle de Calvin-Benson comme la 3-phosphoglycérate. Les réactions produisent également de l'ammoniac (NH 3 ), qui est capable de diffuser à l'extérieur de l'usine, ce qui conduit à une perte d'azote.
- Un résumé très simplifié est:
- 2 glycolate + ATP → 3-phosphoglycérate + dioxyde de carbone + ADP + NH3
La voie de récupération pour les produits de l'activité oxygénase RuBisCO est plus communément connu sous le nomphotorespiration, car il est caractérisé par la consommation d'oxygène dépendant de la lumière et de la libération de dioxyde de carbone.