

Espèce
Saviez-vous ...
Arrangeant une sélection Wikipedia pour les écoles dans le monde en développement sans internet était une initiative de SOS Enfants. Parrainage d'enfants aide les enfants du monde en développement à apprendre aussi.
En biologie , une espèce (pluriel: les espèces) est l'une des unités de base de la classification biologique et un rang taxonomique. Une espèce est souvent définie comme un groupe d' organismes capables de métissage et production fertile progéniture. Bien que dans de nombreux cas, cette définition est suffisante, la difficulté d'espèces est connu comme définissant la problème des espèces. Mesures différentes sont souvent utilisés, comme similitude de l'ADN, la morphologie ou la niche écologique. Présence de traits spécifiques adaptées localement peut subdiviser en espèces " taxons infraspécifiques "tels que (sous-espèce et en botanique autre taxons sont utilisés, tels que variétés, sous-variétés, et formae).
Espèce hypothétique d'avoir les mêmes ancêtres sont placés dans une genre , basée sur les similitudes. La similitude des espèces est jugée sur la base de la comparaison des attributs physiques, en particulier leurs ADN des séquences, le cas échéant. Toutes les espèces sont donnés un nom en deux parties , un «nom binomial". La première partie d'un nom binomial est le nom générique , le genre de l'espèce. La deuxième partie est soit appelé le nom spécifique (un terme utilisé uniquement en zoologie) ou épithète spécifique (le terme utilisé en botanique, qui peut également être utilisé en zoologie). Par exemple, Boa constrictor est l'une des quatre espèces de la genre de Boa. La première partie du nom est capitalisé, et la deuxième partie a une minuscule. Le nom binomial est écrit en italique et souligné lors de l'impression quand manuscrite.
Une définition utilisable du mot «espèce» et des méthodes fiables d'identification des espèces particulières sont indispensables pour affirmer et tester des théories biologiques et pour mesurer la biodiversité , bien que d'autres niveaux taxonomiques telles que les familles peuvent être considérées dans les études à grande échelle. Espèces éteintes connues seulement des fossiles sont généralement difficiles à attribuer les classements taxonomiques précises, ce est pourquoi les niveaux taxonomiques supérieurs tels que les familles sont souvent utilisés pour les études fossiles.
Le nombre total d'espèces non-bactériennes dans le monde a été estimé à 8,7 millions, avec des estimations antérieures allant de deux millions à 100 millions.
La définition de travail des biologistes
Une définition utilisable du mot «espèce» et des méthodes fiables d'identification des espèces particulières est essentielle pour énoncer et tester des théories biologiques et pour mesurer la biodiversité . Traditionnellement, plusieurs exemples d'une espèce proposées doivent être étudiés pour les caractères unificateurs avant qu'il puisse être considéré comme une espèce. Il est généralement difficile de donner classements taxonomiques précises éteints espèces connues seulement des fossiles.
Certains biologistes peuvent voir espèces comme des phénomènes statistiques, par opposition à l'idée traditionnelle, avec une espèce considérée comme une classe d'organismes. Dans ce cas, une espèce est définie comme une lignée évolutive séparément qui forme une seule pool génique. Bien que les propriétés telles que l'ADN-séquences et la morphologie sont utilisés pour aider à séparer lignées étroitement liés, cette définition a des limites floues. Toutefois, la définition exacte du terme «espèce» est encore controversée, en particulier dans les procaryotes, et ce est ce qu'on appelle la problème des espèces. Les biologistes ont proposé une série de définitions plus précises, mais la définition utilisée est un choix pragmatique qui dépend des particularités des espèces préoccupantes.
Noms et espèces communes
Les noms couramment utilisés pour taxons végétaux et animaux correspondent parfois à des espèces: par exemple, " lion "," morse ", et" Camphrier "- chaque référence à une espèce Dans d'autres cas les noms communs ne ont pas: par exemple,." cerf »fait référence à un famille de 34 espèces, y compris De Eld Deer, Red Deer et Elk (Wapiti). Les deux dernières espèces étaient autrefois considérés comme une seule espèce, illustrant la façon dont les limites des espèces peuvent changer avec les connaissances scientifiques accrue.
Placement dans les genres
Idéalement, une espèce est donné, un nom scientifique formelle, bien que dans la pratique il ya de très nombreuses espèces anonymes (qui ne ont été décrites, ne sont pas citées). Lorsqu'une espèce est nommé, il est placé dans un genre . D'un point de vue scientifique, cela peut être considéré comme une hypothèse que l'espèce est plus étroitement lié à d'autres espèces dans son genre (le cas échéant) que les espèces d'autres genres. Espèce et de genre sont généralement définis comme partie d'une hiérarchie taxonomique plus grande. Les rangs taxonomiques les plus connus sont, dans l'ordre: la vie , domaine, royaume, phylum, classe, afin, famille, genre et espèces. Cette affectation à un genre ne est pas immuable; plus tard, un autre (ou le même) taxonomiste peut l'attribuer à un genre différent, dans ce cas, le nom changera également.
Dans la nomenclature biologique, le nom d'une espèce est un nom en deux parties (un nom binomial ), traités comme latine , bien que les racines de ne importe quelle langue peuvent être utilisées ainsi que les noms de lieux ou de personnes. Le nom générique apparaît en premier (avec son leader lettre en majuscule), suivi par un second mandat. La terminologie utilisée pour le second terme diffère entre nomenclatures zoologique et botanique.
- Dans la nomenclature zoologique, la deuxième partie du nom peut être appelé le nom spécifique ou l'épithète spécifique. Par exemple, les loups gris appartiennent à l'espèce Canis lupus, coyotes à Canis latrans, chacals dorés à Canis aureus, etc., et tous ceux qui appartiennent au genre Canis (qui contient également de nombreuses autres espèces). Pour le loup gris, le nom du genre est Canis, du nom spécifique ou épithète spécifique est le lupus et les binôme, le nom de l'espèce, est Canis lupus.
- Dans la nomenclature botanique, la deuxième partie de ce nom ne peut être appelé épithète spécifique. Le «nom spécifique» en botanique est toujours la combinaison du nom de genre et l'épithète spécifique. Par exemple, les espèces communément appelé le pin des marais est Pinus palustris; le nom du genre est Pinus, l'épithète spécifique est palustris, le nom spécifique est Pinus palustris.
Cette convention de nommage binomiale, plus tard, formalisée dans le biologique codes de nomenclature, a été utilisé la première fois par Leonhart Fuchs et présenté comme la norme par Carolus Linnaeus 1753 dans son Species Plantarum (suivies par son Systema Naturae 1758, 10ème édition). A cette époque, la théorie biologique chef était que les espèces représentées actes indépendants de la création de Dieu et ont donc été considérés objectivement réel et immuable, donc l'hypothèse d'une origine commune ne se appliquent pas.
Noms abrégés
Livres et articles parfois intentionnellement ne pas identifier les espèces pleinement et utilisent l'abréviation «sp." dans le "spp." singulier ou au pluriel à la place de l'épithète spécifique:. par exemple, Canis sp Cela se produit généralement dans les types de situations suivantes:
- Les auteurs sont convaincus que certaines personnes appartiennent à un genre particulier, mais ne sont pas sûr de quelle espèce exacte ils appartiennent. Ce est particulièrement fréquente dans la paléontologie .
- Les auteurs utilisent "spp." comme un court façon de dire que quelque chose se applique à de nombreuses espèces dans un genre, mais ne veut pas dire qu'il se applique à toutes les espèces au sein de ce genre. Si les scientifiques signifient que quelque chose se applique à toutes les espèces dans un genre, ils utilisent le nom du genre sans l'épithète spécifique.
Dans les livres et articles, genre et espèce noms sont généralement imprimés en italique. Les abréviations telles que "sp.", "Spp.", "Subsp.", Etc. ne devraient pas être en italique.
Codes d'identification
Plusieurs codes ont été mis au point pour identifier les espèces particulières. Par exemple:
- NCBI emploie un «taxid 'identifiant numérique ou de la taxonomie, un« identifiant unique stable », par exemple, le taxid de H. sapiens est 9606
- KEGG utilise un code à trois lettres pour un nombre limité d'organismes; dans ce code, par exemple, H. sapiens est tout simplement HSA;
- UniProt emploie un «mnémonique organisme" de pas plus de cinq caractères alphanumériques, par exemple humaines pour H. sapiens
Difficulté de définir «espèce» et identifier les espèces particulières

Il est étonnamment difficile de définir le mot «espèces» d'une manière qui se applique à tous les organismes d'origine naturelle, et le débat parmi les biologistes sur la façon de définir les «espèces» et comment identifier les espèces réels est appelé problème des espèces. Plus de deux douzaines définitions distinctes de «espèces» sont en usage chez les biologistes.
La plupart des manuels suivent la définition de Ernst Mayr d'une espèce comme «groupes de réellement ou potentiellement métissage des populations naturelles, qui sont la reproduction isolées à partir d'autres groupes".
Divers éléments de cette définition servent à exclure certains accouplements inhabituelles ou artificiels:
- Ceux qui se produisent seulement en captivité (lorsque les partenaires d'accouplement normales de l'animal ne sont pas disponibles) ou à la suite de l'action humaine délibérée
- Les animaux qui peuvent être physiquement et physiologiquement capable de se accoupler, mais, pour diverses raisons, ne fait pas normalement dans la nature
La définition typique d'école ci-dessus fonctionne bien pour la plupart organismes multicellulaires, mais il ya plusieurs types de situations dans lesquelles il se décompose:
- Par définition, il ne se applique qu'aux organismes qui reproduire sexuellement. Donc, il ne fonctionne pas pour à reproduction asexuée organismes unicellulaires et le nombre relativement faible parthénogénétique ou organismes multicellulaires apomictiques. Le terme " phylotype "est souvent utilisée pour ces organismes.
- Les biologistes ne savent souvent pas si deux groupes morphologiquement similaires d'organismes sont «potentiellement» capable de se reproduire.
- Il existe des variations considérables dans la mesure dans laquelle l'hybridation peut réussir dans des conditions naturelles, ou même dans la mesure dans laquelle certains organismes utilisent la reproduction sexuée entre les individus pour se reproduire.
- En espèces de cycle, les membres des populations adjacentes se croisent avec succès, mais les membres de certaines populations non adjacents ne le font pas.
- Dans quelques cas, il peut être physiquement impossible pour les animaux qui sont membres de la même espèce à accoupler. Cependant, ce sont des cas, comme dans les races de chiens, où l'intervention humaine a provoqué des changements morphologiques brutes, et sont donc exclus par le concept d'espèce biologique.
Transfert horizontal de gènes rend encore plus difficile de définir le mot «espèce». Il ya des preuves solides de transfert horizontal de gènes entre des groupes très différents de les procaryotes, et au moins de temps en temps entre des groupes différents de les eucaryotes ; et Williamson affirme qu'il existe des preuves pour elle dans certains crustacés et échinodermes. Toutes les définitions du mot «espèce» supposent que l'organisme reçoit tous ses gènes d'un ou deux parents qui sont très semblable à cet organisme, mais le transfert horizontal de gènes rend cette hypothèse fausse.
Définitions des espèces
La question de la meilleure façon de définir les «espèces» est celui qui a occupé les biologistes depuis des siècles, et le débat se est devenu connu sous le nom problème des espèces. Darwin a écrit dans le chapitre II de De l'origine des espèces :
- Pas une définition a satisfait tous les naturalistes; pourtant, chaque naturaliste sait vaguement ce qu'il veut dire quand il parle d'une espèce. Généralement, le terme inclut l'élément inconnu d'un acte distinct de la création.
Mais plus tard, en The Descent of Man, se adressant "La question de savoir si l'humanité se compose d'un ou de plusieurs espèces", Darwin a révisé son opinion à-dire:
- ce est un effort désespéré pour décider ce point pour des raisons sonores, jusqu'à une définition du terme «espèce» est généralement admis; et la définition ne doit pas inclure un élément qui ne peut éventuellement être déterminé, comme un acte de création.
La théorie moderne de l'évolution dépend d'une redéfinition fondamentale des «espèces». Avant Darwin, les naturalistes consulté espèces types comme idéales ou générales, qui pourrait être illustré par un spécimen idéal portant tous les traits généraux de l'espèce. Les théories de Darwin détourné l'attention de l'uniformité de la variation et du général au particulier. Selon l'historien intellectuelle Louis Menand,
- Une fois que notre attention est redirigé vers l'individu, nous avons besoin d'une autre manière de faire des généralisations. Nous ne sommes plus intéressés par la conformité d'un individu à un type idéal; nous nous intéressons dans la relation d'un individu aux autres personnes avec lesquelles elle interagit. Pour généraliser sur les groupes de personnes en interaction, nous devons laisser tomber la langue de types et essences, qui est prescriptive (nous dire ce que pinsons devraient être), et d'adopter la langue de statistiques et probabilités, qui est prédictive (ce que nous dit le pinson moyenne, dans des conditions spécifiées, est susceptible de le faire). Relations seront plus importants que les catégories; fonctions, qui sont variables, seront plus importants que fins; transitions seront plus importants que les frontières; séquences seront plus importants que les hiérarchies.
Ce résultats de décalage dans une nouvelle approche de «espèce»; Darwin conclu que les espèces sont ce qu'elles semblent être: des idées, qui sont provisoirement utile pour les groupes de personnes qui interagissent nommer. «Je regarde les espèces terme", écrit-il, "comme un arbitraire donnée pour des raisons de commodité à un ensemble d'individus ressemblant étroitement à l'autre ... Il ne diffère pas essentiellement du mot variété, qui est donné à moins distincte et plusieurs formes fluctuantes. Le terme variété, encore une fois, en comparaison avec de simples différences individuelles, est également appliquées de manière arbitraire, et pour plus de commodité. "
Pratiquement, les biologistes définissent espèces comme les populations d'organismes qui ont un niveau élevé de similitude génétique. Cela peut refléter une adaptation à la même créneau, et le transfert du matériel génétique d'un individu à d'autres, par divers moyens. Le niveau exact de similitude utilisé dans une telle définition est arbitraire, mais ce est la définition la plus couramment utilisée pour les organismes qui se reproduisent de façon asexuée ( reproduction asexuée), tels que des plantes et micro-organismes.
Cette absence de tout concept d'espèce clair microbiologie a conduit certains auteurs faisant valoir que le terme «espèce» ne est pas utile lorsque l'on étudie l'évolution bactérienne. Au lieu de cela ils voient gènes se déplacer librement entre les bactéries, même lointainement apparentées, avec l'ensemble du domaine bactérienne étant une piscine d'un seul gène. Néanmoins, une sorte de règle de base a été établi, en disant que les espèces de bactéries ou Archaea avec ARNr 16S séquences géniques similaires de 97% à l'autre doivent être vérifiées par ADN-hybridation de l'ADN si elles appartiennent à la même espèce ou non. Ce concept a été mis à jour récemment, disant que la frontière des 97% était trop faible et peut être portée à 98,7%.
Dans l'étude des organismes à reproduction sexuée, où le matériel génétique est partagée à travers le processus de la reproduction, la capacité des deux organismes de se croiser et de produire une descendance fertile des deux sexes est généralement acceptée comme un indicateur simple que les organismes partagent suffisamment gènes à prendre en considération les membres de la même espèce. Ainsi une «espèce» est un groupe d'organismes métissage.
Cette définition peut être étendue à dire qu'une espèce est un groupe d'organismes qui pourraient se croiser - poissons pourrait encore être classé comme la même espèce, même se ils vivent dans différents lacs, tant qu'ils pouvaient encore se croiser ont-ils jamais d'entrer en communiquer les uns avec les autres. D'autre part, il existe de nombreux exemples de séries de trois ou plusieurs populations distinctes, où les individus de la population au milieu peuvent se croiser avec les populations de chaque côté, mais les individus des populations des deux côtés ne peuvent pas se croiser. Ainsi, on pourrait dire que ces populations constituent une seule espèce, ou de deux espèces distinctes. Ce ne est pas un paradoxe; ce est la preuve que les espèces sont définies par la fréquence des gènes, et ont donc des limites floues.
Par conséquent, une définition unique, universel de «espèce» est nécessairement arbitraire. Au lieu de cela, les biologistes ont proposé une série de définitions; dont la définition d'une biologistes utilise est un choix pragmatique, en fonction des particularités de la recherche de ce que biologiste.
Dans la pratique, ces définitions coïncident souvent, et les différences entre eux sont plus une question de l'accent que la contradiction pure et simple. Néanmoins, aucun concept d'espèce encore proposé est entièrement objective, ou peut être appliqué dans tous les cas sans recourir au jugement. Compte tenu de la complexité de la vie, certains ont fait valoir qu'une telle définition objective est, selon toute vraisemblance impossible, et les biologistes doit se contenter de la définition la plus pratique.
Pour la plupart des vertébrés , ce est le concept de l'espèce biologique (BSC), et dans une moindre mesure (ou à des fins différentes) le concept d'espèce phylogénétique (CFP). Beaucoup BSC sous-espèces sont considérées comme des espèces en vertu de la CFP; la différence entre le BSC et la CFP peut se résumer dans la mesure où ce que le BSC définit une espèce à la suite de l'histoire de l'évolution manifeste, alors que la CFP définit une espèce comme une conséquence de potentiel évolutif manifeste. Ainsi, une espèce de la CFP est "fait" dès qu'une lignée évolutive a commencé à se séparer, alors qu'une espèce de BSC commence à exister que lorsque la séparation de la lignée est terminée. En conséquence, il peut y avoir conflit considérable entre classifications alternatives basées sur la CFP par rapport BSC, car ils diffèrent complètement dans leur traitement des taxons qui serait considérée comme sous-espèces dans le cadre du dernier modèle (par exemple, les nombreuses sous-espèces de abeilles ).
Espèces typologiques
Un groupe d'organismes dans lesquels les individus sont membres de l'espèce se ils sont conformes suffisamment pour certaines propriétés fixes. Les grappes de variations ou des phénotypes au sein de spécimens (ce est à dire plus ou queues courtes) seraient différencier les espèces. Cette méthode a été utilisée comme une méthode "classique" de la détermination espèces, comme avec Linnaeus au début de la théorie de l'évolution. Cependant, nous savons maintenant que différents phénotypes ne constituent pas toujours espèces différentes (par exemple: un quatre ailes Drosophila né d'une mère deux ailes ne est pas une espèce différente). Espèces citées dans cette manière sont appelés morphoespèces ou espèce nominale.
Espèces évolutionnaires
Une seule lignée évolutive des organismes au sein de laquelle les gènes peut être partagée, et qui maintient son intégrité par rapport à d'autres lignées à la fois temps et l'espace. À un certain moment dans l'évolution d'un tel groupe, certains membres peuvent se écarter de la population principale et évoluer vers une sous-espèce, un processus qui peut éventuellement conduire à la formation d'une nouvelle espèce si l'isolement (géographique ou écologique) est maintenue. Une espèce qui donne lieu à une autre espèce est une espèce paraphyletic, ou paraspecies.
(Cladistiques) espèces phylogénétiques
Un groupe d'organismes qui part un ancêtre; une lignée qui conserve son intégrité par rapport à d'autres lignées à la fois temps et l'espace. À un certain point dans la progression d'un tel groupe, les membres peuvent se écarter de l'autre: quand une telle divergence devient suffisamment claire, les deux populations sont considérées comme des espèces distinctes. Ceci diffère de l'évolution des espèces en ce que les espèces parentes va taxonomique éteinte quand une nouvelle espèce évolue, les populations de mère et la fille qui fait maintenant deux nouvelles espèces. La sous-espèce en tant que tels ne sont pas reconnus dans cette approche; soit une population est une espèce phylogénétique ou il ne est pas taxonomique distinguer.
Autre
- Espèces écologiques
- Un ensemble d'organismes adaptés à un ensemble particulier de ressources, appelé une niche, dans l'environnement. Selon ce concept, les populations forment des grappes de phénétiques discrètes que nous reconnaissons comme des espèces parce que les processus écologiques et évolutifs qui contrôlent la façon dont les ressources sont réparties ont tendance à produire ces clusters.
- Espèces biologiques / reproduction
- Deux organismes qui sont en mesure de se reproduire naturellement à produire une descendance fertile des deux sexes. Organismes qui peuvent se reproduire, mais presque toujours font hybrides stériles d'au moins un sexe, comme un mule, bardot ou F1 mâle cattalo ne sont pas considérés être les mêmes espèces.
- Espèces biologiques / isolement
- Un ensemble de populations réellement ou potentiellement métissage. Ce est généralement une formulation utile pour les scientifiques travaillant avec des exemples de l'taxons supérieurs comme les mammifères, les poissons et les oiseaux vivants, mais plus problématique pour les organismes qui ne se reproduisent pas sexuellement. Les résultats des expériences de reproduction effectuées dans des conditions artificielles peuvent ou peuvent ne pas refléter ce qui se passerait si les mêmes organismes rencontrés l'autre à l'état sauvage, ce qui rend difficile d'évaluer si oui ou non les résultats de ces expériences sont significatives en référence à des populations naturelles.
- Espèces génétiques
- Basé sur la similarité de l'ADN des individus ou des populations. Techniques de comparer la similitude des ADN comprend Hybridation de l'ADN, et l'empreinte génétique (ou Codes-barres ADN).
- espèces de cohésion
- La population la plus inclusive des individus ayant le potentiel pour la cohésion phénotypique par des mécanismes de cohésion intrinsèques. Ce est une extension du concept d'espèce mate-de reconnaissance pour permettre mécanismes d'isolement après l'accouplement; peu importe si les populations peuvent se hybrider avec succès, ils sont encore des espèces distinctes de cohésion si le montant de l'hybridation est insuffisante pour mélanger complètement leur respective pools génétiques.
- Evolutionarily Unité significative (ESU)
- Une évolutif unité significative est une population d'organismes qui est considéré comme distinct à des fins de conservation. Souvent désigné comme une espèce ou d'une espèce sauvage, une UES a également plusieurs définitions possibles, qui coïncident avec les définitions des espèces.
- Espèces phénétiques
- Basé sur phenetics.
| Chercher micro-espèces dans Wiktionnaire, le dictionnaire libre. |
- Micro-espèces
- Une espèce avec très peu de variabilité génétique, généralement celui qui se reproduit par apomixie.
- Espèces de reconnaissance
- Basé sur le système reproducteur partagés, y compris le comportement d'accouplement. Le concept de reconnaissance des espèces a été introduit par Hugh EH Paterson, après le travail plus tôt par Wilhelm Petersen.
- Espèces Mate-reconnaissance
- Un groupe d'organismes qui sont connus de reconnaître l'autre comme des partenaires potentiels. Comme le concept d'espèce d'isolement ci-dessus, il ne se applique qu'aux organismes qui se reproduisent sexuellement. Contrairement au concept d'espèce d'isolement, il se concentre spécifiquement sur l'isolement reproductif avant l'accouplement.
Nombre d'espèces
Ayant à l'esprit les problèmes mentionnés ci-dessus avec des espèces catégorisation, les numéros suivants sont seulement un guide. Sur la base de diverses discussions de la première décennie du nouveau millénaire, les comptes peuvent être grossièrement décomposent comme suit:
Nombre de espèces procaryotes, les bactéries Domain
- Ce nombre est très difficile à évaluer, mais la gamme discuté varie de dizaines de milliers de milliards; la plupart des approches et des études récentes semblent favoriser le nombre de plus grande ampleur. Un plus petit nombre se posent des hypothèses fondées sur un plafonnement de l'identification de nouvelles espèces (qui a des explications techniques autres que ce que moins d'espèces restent à être identifiés). Un plus grand nombre abordent le fait que le succès de culture de bactéries n'a été atteint que dans la moitié des Eubacteria identifié (où le manque de réussite dans les tentatives à la culture un bactériennes limites isoler capacités pour étudier et délimiter de nouvelles espèces), et face à la difficulté d'appliquer botanique traditionnelle et les définitions zoologiques d'espèces à reproduction asexuée à bactéries (où plus séquençage moderne et approches moléculaires soutiennent espèces supérieures de décompte).
Nombre de espèces procaryotes, Domain Archaea
- Comme autre domaine microbienne, les enjeux et les difficultés de bactéries domaine portent également à tout comptage des espèces de Archaea, d'autant plus compte tenu de leur différents habitats extrêmes. Comme l'article de Wikipedia sur le Archéobactéries notes (Classification, l'article espèces), la «classification des archées en espèces est également controversée», comme ils se reproduisent aussi asexuée (même éliminer applicabilité des espèces définitions basées sur le croisement), et font face aux mêmes difficultés liées à l'isolement de l'organisme et la culture (voir citations pour les bactéries, ci-dessus). Archaebacteria ont été montré pour présenter des taux élevés de transfert horizontal de gènes (résultant d'un apparenté bactérienne du sexe), y compris entre les organismes bien distincts basés sur l'analyse génomique. Comme le Article Archaea notes, «[c] connaissance dusyst sur la diversité génétique est fragmentaire et le nombre total d'espèces archéens ne peut être estimé avec précision" ... si comme les bactéries de domaine, le nombre de cultivé et étudié phylums par rapport au total est faible (à partir de 2005, moins de 50% des phylums connus de culture). Pris dans leur ensemble, un très grand nombre de types archéobactériennes uniques sont susceptibles, comme dans le cas des bactéries domaine.
Nombre d' eucaryotes espèces
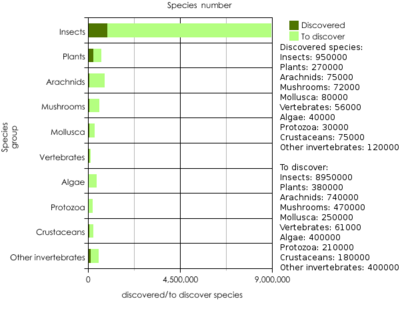
Ce nombre a historiquement varie de quelques millions à environ 100 millions. Cependant ces chiffres élevés, qui étaient fondées sur la diversité potentielle de marine et arthropodes profonde, sont maintenant considérés comme peu probable. Le nombre total d'espèces eucaryotes est susceptible d'être 5 ± 3.000.000 dont environ 1,5 millions ont été déjà nommé.
Certaines estimations âgés, pour diverses eucaryote phylums sont (y compris quelques chiffres mis à jour):
- Jusqu'à 1,5 million de champignons ;
- 3067 algues brunes
- 17 000 lichens ;
- 321 212 plantes , y compris:
- 1367555 non-insectes animaux , y compris:
- 1305250 invertébrés
- 2175 coraux
- 85000 mollusques
- Autant que 1,1 millions arachnides, y compris ~ 1000000 les acariens et d'autres arachnides ~ 100000
- 47000 crustacés
- 68 827 autres invertébrés;
- 63 649 vertébrés
- 31 300 poissons ,
- 7093 amphibiens,
- 9768 reptiles ,
- 9998 oiseaux ,
- 5490 mammifères ;
- 1305250 invertébrés
- Autant que 10-30000000 insectes ;
À l'heure actuelle, des organisations telles que la Initiative taxonomique mondiale, le Institut européen de Taxonomie et Census of Marine Life (ce dernier uniquement pour les organismes marins) se efforcent d'améliorer la taxonomie et mettre en œuvre espèces jusqu'ici inconnu avec un système de taxonomie, bien que les connaissances actuelles ne couvre qu'une partie des organismes de la biosphère et donc ne permet pas une compréhension complète de la fonctionnement de l'environnement. L'humanité est également un essorage actuellement hors espèces non découvertes à un rythme sans précédent, ce qui signifie que, même avant une nouvelle espèce a eu la chance d'être étudiés et classés, il peut déjà avoir disparu.
Importance dans la classification biologique
L'idée des espèces a une longue histoire. Ce est un des niveaux les plus importants de classification, pour plusieurs raisons:
- Il correspond souvent à ce que les laïcs traitent comme les différents types de base de l'organisme - les chiens sont une espèce, chats autre.
- Ce est la norme nomenclature binomiale (ou nomenclature trinôme) par lequel les scientifiques se réfèrent généralement à des organismes.
- Ce est le niveau taxonomique le plus élevé qui ne peuvent pas être rendue plus ou moins d'inclusion.
Après des années d'utilisation, le concept demeure au centre de la biologie et une foule de domaines connexes, et pourtant reste aussi parfois mal définies.
Conséquences de la cession de l'état des espèces
La désignation d'une espèce particulière peut être considérée comme une hypothèse sur les relations évolutives et la distinction de ce groupe d'organismes. Comme plus d'informations tombe sous la main, l'hypothèse peut être confirmée ou infirmée. Parfois, surtout dans le passé lorsque la communication a été plus difficile, les taxonomistes qui travaillent dans l'isolement ont donné deux noms distincts pour les organismes individuels tard identifiés comme les mêmes espèces. Lorsque deux espèces citées sont découverts à être de la même espèce, le nom de l'espèce est généralement plus retenu, et le nom de l'espèce nouvelle a chuté, un processus appelé synonymization, ou familièrement, comme la formation de grumeaux. Division d'un taxon en plusieurs, souvent nouveaux, taxons est appelé fractionnement. Les taxonomistes sont souvent appelés «débardeurs» ou «diviseurs» par leurs collègues, en fonction de leur approche personnelle à reconnaître les différences ou les points communs entre les organismes (voir débardeurs et les répartiteurs).
Traditionnellement, les chercheurs se appuyaient sur les observations des différences anatomiques et sur des observations de savoir si les différentes populations ont pu se croiser avec succès, de distinguer les espèces; l'anatomie et le comportement de reproduction sont toujours importantes pour la désignation du statut des espèces. En conséquence de la révolutionnaire (et toujours en cours) avance dans les techniques de recherche microbiologiques, y compris l'analyse de l'ADN, dans les dernières décennies, beaucoup de connaissances sur les différences et les similitudes entre les espèces est devenu disponible. Beaucoup de populations autrefois considérés comme des espèces distinctes sont désormais considérés comme un seul taxon, et de nombreuses populations anciennement regroupées ont été divisés. Toute niveau taxonomique (espèce, genre, famille, etc.) peut être synonymie ou de scission, et aux niveaux taxonomiques élevés, ces révisions ont été encore plus profonde.
De point de vue taxonomique, les groupes au sein d'une espèce peut être définie comme étant d'un taxon hiérarchiquement inférieure à une espèce. En zoologie seulement le sous-espèce est utilisé, tandis que dans la botanique du variété, sous-variété, et forme sont utilisés ainsi. En biologie de la conservation, la notion de unités évolutionnaires significatives (ESU) est utilisé, qui peut définir deux espèces ou plus petits segments de population distincts. Identifier et nommer les espèces est la providence de alpha taxonomie.
Le développement historique de la notion de l'espèce


Dans les premières œuvres de la science, une espèce était tout simplement un organisme individuel qui représentait un groupe d'organismes similaires ou presque identiques. Pas d'autres relations au-delà de ce groupe ont été impliqués. Aristote a utilisé les mots genre et espèces pour signifier catégories génériques et spécifiques. Aristote et d'autres scientifiques pré-darwiniens ont pris l'espèce doit être distinct et immuable, avec une " essence ", comme les éléments chimiques . Lorsque les premiers observateurs ont commencé à développer des systèmes d'organisation pour les êtres vivants, ils ont commencé à placer des espèces autrefois isolées dans un contexte. Beaucoup de ces régimes de délimitation début serait maintenant considéré comme fantaisiste et ces consanguinité inclus fondée sur la couleur (toutes les plantes à fleurs jaunes) ou de comportement (serpents, scorpions et certaines fourmis piqueuses).
John Ray (1686), un naturaliste anglais, était le premier à donner une définition biologique de l'espèce à long terme.
Dans le scientifique suédois du 18ème siècle Carolus Linnaeus classé organismes en fonction des différences dans la forme de appareil reproducteur. Bien que son système de classification trie organismes selon leur degré de similitude, il fait aucune réclamation au sujet de la relation entre les espèces semblables. A cette époque, il était encore largement admis qu'il n'y avait pas de lien organique entre espèces, peu importe comment ils sont apparus semblables. Cette approche a également suggéré un type de l'idéalisme: la notion que chaque espèce existait comme une «forme idéale". Bien qu'il existe toujours des différences (même si parfois minute) entre les organismes individuels, Linnaeus considéré telle variation problématique. Il se efforçait d'identifier les organismes individuels qui étaient exemplaire de l'espèce, et a examiné d'autres organismes non exemplaires comme déviants et imparfaite.
Dès le 19e siècle, la plupart des naturalistes compris que les espèces pourraient changer de forme au fil du temps, et que l'histoire de la planète prévu suffisamment de temps pour des changements majeurs. Jean-Baptiste Lamarck, dans sa Philosophie zoologique 1809, a offert l'un des premiers arguments logiques contre le créationnisme . Le nouveau accent était mis sur la détermination de la façon dont une espèce pourrait changer au fil du temps. Lamarck a suggéré qu'un organisme pourrait passer sur un caractère acquis à sa descendance, ce est à dire, la girafe de long cou a été attribuée à des générations de girafes d'étirement pour atteindre les feuilles de la cime des arbres plus élevés (cet exemple bien connu et simpliste, cependant, ne est pas rendre justice à l'étendue et la subtilité des idées de Lamarck). Avec l'acceptation de la sélection naturelle idée de Charles Darwin dans les années 1860, cependant, le point de vue de l'évolution de Lamarck orientée vers un but, également connu comme un processus téléologique, a été éclipsé. L'intérêt récent pour hérédité des caractères acquis se articule autour processus épigénétiques, par exemple, méthylation, qui ne affecte pas les séquences d'ADN, mais au lieu de modifier l'expression d'une manière héréditaire. Ainsi, Neo-lamarckisme, comme il est parfois appelé, ne est pas un défi à la théorie de l'évolution par la sélection naturelle.
Charles Darwin et Alfred Wallace fourni ce que les scientifiques considèrent désormais comme le plus puissant et convaincant théorie de l'évolution . Darwin a fait valoir qu'il était populations qui ont évolué, et non des individus. Son argument invoqué un changement radical de perspective de celle de Linné: plutôt que de définir les espèces en termes idéales (et la recherche d'un représentant idéal et en rejetant écarts), Darwin considéré variation entre les individus d'être naturel.Il a fait valoir en outre que la variation, loin d'être problématique, fournit en fait l'explicationde l'existence d'espèces distinctes.
Les travaux de Darwin a attiré sur Thomas Malthus l'intuition de ce que le taux de croissance d'une population biologique sera toujours dépasser le taux de croissance des ressources de l'environnement, tels que l'approvisionnement alimentaire. En conséquence, Darwin a fait valoir, ne sont pas tous les membres d'une population seront en mesure de survivre et se reproduire. Ceux qui fait la volonté, en moyenne, ceux possédant des variations-même minimes-qui les rendent un peu mieux adaptés à l'environnement. Si ces traits variables sont héréditaires, puis les descendants des survivants sera également posséder eux. Ainsi, au fil des générations, les variations adaptatives seront accumuler dans la population, tandis que les traits de contre-adaptative auront tendance à être éliminé.
Si une modification est adaptatif ou non adaptatif dépend de l'environnement: des environnements différents favorisent différents traits. Depuis l'environnement sélectionne efficacement, ce qui organismes vivent à reproduire, il est l'environnement (la «lutte pour l'existence») qui sélectionne les traits d'être transmis. Telle est la théorie de l'évolution par la sélection naturelle . Dans ce modèle, la longueur du cou d'une girafe serait expliqué en posant que les proto-girafes avec un cou plus long auraient eu un avantage reproductif important pour ceux qui cous plus courts. Au fil des générations, la population entière serait une espèce d'animaux au long cou.
En 1859, quand Darwin a publié sa théorie de la sélection naturelle, le mécanisme derrière l'héritage de traits individuels était inconnu. Bien que Darwin a fait quelques spéculations sur la façon dont les traits sont hérités ( pangenèse), sa théorie ne repose que sur le fait que les traits héréditaires existent , et sont variables (ce qui rend son exploit encore plus remarquable.) Bien que le document de Gregor Mendel sur la génétique a été publié en 1866, son importance n'a pas été reconnue. Il a fallu attendre 1900 que son travail a été redécouvert par Hugo de Vries, Carl Correns et Erich von Tschermak, qui ont réalisé que les "les traits héréditaires» dans la théorie de Darwin sont gènes.
La théorie de l'évolution des espèces par la sélection naturelle a deux implications importantes pour les discussions d'espèces de conséquences qui remettent fondamentalement les hypothèses sous-jacentes de Linné la taxonomie. Tout d'abord, il suggère que les espèces ne sont pas seulement similaire, ils peuvent en fait être liés. Certains étudiants de Darwin font valoir que tous les espèces sont les descendants d'un ancêtre commun. Deuxièmement, elle suppose que «espèce» ne sont pas homogènes, fixes, les choses permanentes; membres d'une espèce sont tous différents, et au fil du temps le changement de l'espèce. Ceci suggère que les espèces ne sont pas des limites claires, mais sont plutôt des effets statistiques momentanées de l'évolution constante de gènes fréquences. On peut toujours utiliser la taxonomie de Linné pour identifier les plantes et les animaux individuels, mais on ne peut plus penser à des espèces comme indépendants et immuables.
La montée d'une nouvelle espèce à partir d'une lignée parentale est appelé spéciation.Il n'y a aucune ligne claire délimitation des espèces ancestrales des espèces descendants.
Bien que la compréhension scientifique actuelle des espèces suggère qu'il n'y a pas de manière rigoureuse et complète de distinguer entre les différentes espèces dans tous les cas, les biologistes continuent à rechercher des moyens concrets pour opérationnaliser l'idée. Une des définitions biologiques les plus populaires des espèces est en termes d'isolement reproductif; si deux créatures ne peuvent pas se reproduire pour produire une descendance fertile des deux sexes, alors ils sont dans différentes espèces. Cette définition englobe un certain nombre de limites d'espèces intuitives, mais il reste imparfaite. Il n'a rien à dire sur les espèces qui se reproduisent de façon asexuée, par exemple, et il est très difficile à appliquer à des espèces éteintes. En outre, les frontières entre espèces sont souvent floue: il existe des exemples où les membres d'une population peuvent produire une descendance fertile des deux sexes avec une deuxième population, et les membres de la deuxième population peuvent produire une descendance fertile des deux sexes avec les membres d'une troisième population, mais les membres de la première et la troisième population ne peuvent pas produire une descendance fertile, ou ne peuvent produire une descendance fertile du sexe homozygote. Par conséquent, certaines personnes rejettent cette définition de l'espèce.
Richard Dawkins définit deux organismes comme conspécifiques si et seulement si ils ont le même nombre de chromosomes et, pour chaque chromosome, les deux organismes ont le même nombre de nucléotides ( L'Horloger aveugle , p. 118). Cependant, la plupart sinon tous les taxonomistes seraient en désaccord. Par exemple, dans de nombreux amphibiens, notamment en Nouvelle-Zélande Leiopelma les grenouilles, le génome est constitué de chromosomes «de base» qui sont la plupart du temps chromosomes invariables et accessoires, dont existent un certain nombre de combinaisons possibles. Même si les nombres chromosomiques sont très variables entre les populations, celles-ci peuvent se croiser avec succès et forment une unité évolutive unique. Chez les plantes, polyploïdie est extrêmement courante avec peu de restrictions sur métissage; comme des individus ayant un nombre impair de jeux de chromosomes sont généralement stériles, en fonction du nombre réel de jeux de chromosomes présent, cela se traduit dans la situation étrange où des individus de la même unité d'évolution peuvent se croiser avec certains autres et certains ne peuvent pas, avec toutes les populations étant éventuellement liée à former un pool génétique commun.
La classification des espèces a été profondément affecté par les progrès technologiques qui ont permis aux chercheurs de déterminer la parenté fondée sur des marqueurs moléculaires, en commençant par les relativement brut sanguins tests de précipitation de plasma dans le milieu du 20e siècle à révolutionnaires de Charles Sibley les études d'hybridation ADN-ADN dans 1970 conduisant à des techniques de séquençage de l'ADN. Les résultats de ces techniques ont causé des changements révolutionnaires dans les catégories taxonomiques supérieurs (tels que embranchements et classes), résultant de la réorganisation de nombreuses branches de l' arbre phylogénétique ( voir aussi: phylogénie moléculaire). Pour les catégories taxonomiques ci-dessous genres , les résultats ont été mitigés jusqu'à présent; le rythme du changement évolutif au niveau moléculaire est plutôt lent, ce qui donne des différences claires seulement après de longues périodes de séparation de reproduction. résultats d'hybridation ADN-ADN ont conduit à des conclusions erronées, l' Labbe pomarin - Grande phénomène Skua étant un exemple célèbre. Tortues ont été déterminés à évoluer avec seulement un huitième de la vitesse d'autres reptiles au niveau moléculaire, et le taux de moléculaire évolution des albatros est la moitié de ce qui se trouve dans le lieu étroitement lié pétrels. La technique d'hybridation est maintenant obsolète et est remplacée par des approches computationnelles plus fiables pour la comparaison de séquences. Taxonomie moléculaire est pas directement basée sur les processus évolutifs, mais plutôt sur le changement global apporté sur ces processus. Les processus qui conduisent à la génération et la maintenance de variation tels que mutation, croisement et de sélection ne sont pas uniformes (voir aussi horloge moléculaire). ADN est seulement extrêmement rarement une cible directe de la sélection naturelle plutôt que des changements dans la séquence d'ADN durables au fil des générations étant le résultat de ce dernier; par exemple, silencieuses transition- combinaisons de transversion changerait le point de fusion de la séquence d'ADN, mais pas la séquence des protéines codées et sont donc un exemple possible lorsque, par exemple dans des micro-organismes, une mutation conférant un changement de remise en forme par lui-même.