

Seleção natural
Você sabia ...
Crianças SOS, que corre cerca de 200 sos escolas no mundo em desenvolvimento, organizado esta selecção. Visite o site da SOS Children at http://www.soschildren.org/
A selecção natural é o processo pelo qual favorável transmissível traços se tornam mais comuns em sucessivas gerações de um população de reprodução de organismos e características hereditárias desfavoráveis tornam-se menos comum, devido ao diferencial de reprodução genótipos. A seleção natural atua sobre o fenótipo, ou as características observáveis de um organismo, de tal forma que indivíduos com fenótipos favoráveis têm maior probabilidade de sobreviver e reproduzir do que aqueles com fenótipos menos favoráveis. O fenótipo genético base, genótipo associada com o fenótipo favorável, irá aumentar em freqüência sobre as gerações seguintes. Ao longo do tempo, este processo pode resultar em adaptações que se especializam organismos para especial nichos ecológicos e podem, eventualmente, resultar na surgimento de novas espécies. Em outras palavras, a selecção natural é o mecanismo pelo qual a evolução pode ocorrer em uma população de um organismo específico.
A seleção natural é um dos pilares da moderna biologia . O termo foi introduzido por Charles Darwin em seu inovador livro de 1859 A Origem das Espécies em que a seleção natural foi descrita por analogia com seleção artificial, um processo pelo qual os animais com características consideradas desejáveis por criadores humanos são sistematicamente favorecido para a reprodução. O conceito de selecção natural foi originalmente desenvolvida na ausência de uma teoria válida de herança; no momento da escrita de Darwin, nada se sabia da moderna genética . Embora Gregor Mendel, o pai da genética moderna, foi contemporâneo de Darwin, o seu trabalho estaria na obscuridade até o início do século 20. A união da tradicional A evolução darwinista com descobertas posteriores na clássica e genética molecular é denominada síntese evolutiva moderna. Embora outros mecanismos de evolução molecular, tais como o teoria neutra avançado por Motoo Kimura, foram identificadas como importantes causas de diversidade genética, a seleção natural continua a ser a principal explicação única para a evolução adaptativa.
Princípios gerais

A seleção natural atua sobre um organismo de fenótipo, ou as características físicas. Fenótipo é determinado pela composição genética de um organismo ( genótipo) e o ambiente em que o organismo vive. Muitas vezes, a seleção natural atua sobre específico traços de um indivíduo, e os termos fenótipo e genótipo são usados estritamente para indicar essas características específicas.
Quando os organismos diferentes em uma população possuem diferentes versões de um gene para uma determinada característica, cada uma dessas versões é conhecido como um alelo. É esta variação genética que está por trás traços fenotípicos. Um exemplo típico é que certas combinações de genes para cor dos olhos nos seres humanos, que, por exemplo, dar origem ao fenótipo de olhos azuis. (Por outro lado, quando todos os organismos em uma parcela da população o mesmo alelo para um traço particular, e este estado é estável ao longo do tempo, o alelo é dito ser fixo nessa população.)
Algumas características são reguladas por um único gene, mas a maioria das características são influenciadas pelas interacções de muitos genes. Uma variação em um dos muitos genes que contribui para uma característica pode ter apenas um pequeno efeito sobre o fenótipo; juntos, esses genes podem produzir um contínuo de possíveis valores fenotípicos.
Nomenclatura e uso
O termo "seleção natural" tem um pouco diferentes definições em diferentes contextos. Em termos simples, a "seleção natural" é mais freqüentemente definido para operar em traços hereditários, mas às vezes pode se referir ao sucesso reprodutivo diferencial dos fenótipos independentemente de esses fenótipos são hereditárias. A selecção natural é "cega" no sentido em que o nível de sucesso reprodutivo dos indivíduos é uma função do fenótipo e não de se ou em que extensão é que fenótipo hereditário. Seguindo uso primário de Darwin, o termo é usado frequentemente para se referir tanto a conseqüência de seleção cego e seus mecanismos. Às vezes é útil para distinguir explicitamente entre os mecanismos de selecção e seus efeitos; quando esta distinção é importante, os cientistas definem "seleção natural" especificamente como "aqueles mecanismos que contribuem para a seleção de indivíduos que se reproduzem", sem levar em conta se a base da seleção é hereditária. Este é por vezes referido como "seleção natural fenotípica. '
Traços que causam maior sucesso reprodutivo de um organismo são disse a ser selecionado para a passo que aqueles que reduzem sucesso são selecionados contra. A selecção para uma característica também pode resultar na selecção de outros traços correlacionados que não influenciem directamente a aptidão si. Isto pode ocorrer como resultado de pleiotropy ou ligação do gene.
Fitness
O conceito de fitness é central para a seleção natural. No entanto, como com a seleção natural acima, não é grave divergência de opinião sobre o significado preciso do termo, e Richard Dawkins gere em seus livros posteriores para evitá-lo inteiramente. (Ele dedica um capítulo do seu o fenótipo estendido para discutir os vários sentidos, em que o termo é utilizado.) Embora a aptidão é por vezes coloquialmente entendida como uma qualidade que promove a sobrevivência de um indivíduo em particular - como se ilustra na frase conhecida sobrevivência do mais apto - moderna teoria da evolução define aptidão em termos de reprodução individual. A base dessa abordagem é: se um organismo vive metade do tempo que outros de sua espécie, mas tem o dobro prole sobreviver até a idade adulta produtiva, seus genes se tornarão mais comuns na população adulta da próxima geração. Isto é conhecido como a reprodução diferencial.
Embora a seleção natural age sobre os indivíduos, o seu efeito médio em todos os indivíduos com um genótipo específico corresponde à aptidão de que o genótipo. Muito genótipos de baixa aptidão fazer com que seus portadores a ter poucas ou nenhuma descendência, em média; exemplos incluem muitos humana doenças genéticas como fibrose cística . Condições como a anemia falciforme pode ter baixa aptidão na população humana em geral, mas porque ela confere imunidade contra a malária, tem alto valor de fitness em populações que têm altas taxas de infecção de malária. De um modo geral, a aptidão de um organismo é uma função de aptidões dos seus alelos. Desde aptidão é uma quantidade média, no entanto, é possível uma mutação favorável pode surgir em um indivíduo que não chegam à idade adulta, por razões não relacionadas.
Tipos de selecção
A seleção natural pode atuar em qualquer característica fenotípica, e pressão seletiva pode ser produzido por qualquer aspecto do ambiente, incluindo companheiros e membros da mesma espécie, ou membros de uma mesma espécie. No entanto, isso não implica que a seleção natural é sempre direcional e resulta em evolução adaptativa; seleção natural muitas vezes resulta na manutenção do status quo, eliminando variantes menos aptos.
O unidade de selecção pode ser o indivíduo ou pode ser um outro nível dentro da hierarquia de organização biológica, tais como os genes, as células, e grupos de parentesco. Ainda há debate sobre se a seleção natural age no nível de grupos ou espécies para produzir adaptações que beneficiam um grupo maior, não-parentes. Selecção a um nível diferente, tal como o gene pode resultar num aumento na aptidão para esse gene, e, ao mesmo tempo, reduzir a aptidão dos indivíduos portadores do gene que, num processo denominado intragenômica conflito. No geral, o efeito combinado de todas as pressões de selecção em vários níveis determina a aptidão geral de um indivíduo e, portanto, o resultado da selecção natural.
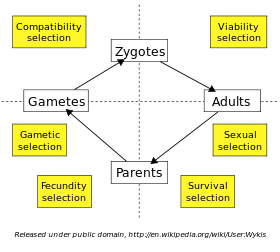
A selecção natural ocorre em todas as fases da vida de um indivíduo. Um organismo individual deve sobreviver até a idade adulta antes que ele possa se reproduzir, e seleção daqueles que chegar a esta fase é chamado de seleção viabilidade. Em muitas espécies, os adultos devem competir uns com os outros para companheiros via seleção sexual, e sucesso nesta competição determina quem vai ser parent a próxima geração. Quando os indivíduos podem se reproduzir mais de uma vez, a maior sobrevida na fase reprodutiva aumenta o número de filhos, chamado de seleção sobrevivência. O fecundidade de ambos os sexos (por exemplo, gigante esperma em certas espécies de Drosophila) pode ser limitada através de seleção fecundidade. A viabilidade das produzido gâmetas podem ser diferentes, enquanto intragenômica conflitos, tais como desvio de segregação entre a gametas haplóides pode resultar na seleção gametic ou gênica. Por fim, a união de algumas combinações de óvulos e espermatozóides podem ser mais compatíveis do que outros; este é denominado selecção compatibilidade.
A seleção sexual
Também é útil fazer uma distinção entre mecanicista seleção ecológica e mais estreito prazo seleção sexual. seleção ecológica abrange qualquer mecanismo de seleção como resultado do ambiente (incluindo parentes, por exemplo, seleção de parentesco, e membros da mesma espécie, por exemplo, competição, infanticídio), enquanto a seleção sexual refere-se especificamente à concorrência entre membros da mesma espécie para companheiros. A seleção sexual pode ser intrasexual, como nos casos de concorrência entre indivíduos do mesmo sexo em uma população, ou intersexual, como nos casos em que um sexo controla o acesso reprodutivo, escolhendo entre uma população de companheiros disponíveis. Mais comumente, intrasexual selecção envolve a competição macho-macho e selecção intersexual envolve a escolha da fêmea de machos adequados, devido à geralmente maior investimento de recursos para uma fêmea de um macho num organismo prole. No entanto, algumas espécies apresentam sexo-papel comportamento revertida em que é do sexo masculino que são mais seletivos na escolha do companheiro; os exemplos mais conhecidos deste padrão ocorrer em alguns peixes da família Syngnathidae, embora susceptíveis exemplos também foram encontrados em anfíbios e aves espécies. Alguns recursos que estão confinados a um sexo somente de uma espécie em particular pode ser explicada pela seleção exercida pelo outro sexo na escolha de um companheiro, por exemplo, a plumagem extravagante de alguns pássaros masculinos. Da mesma forma, a agressão entre os membros do mesmo sexo é por vezes associada com características muito distintas, como os chifres de veados, que são usados em combate com outros veados. Mais geralmente, a selecção intrasexual é muitas vezes associada com dimorfismo sexual, incluindo diferenças no tamanho corporal entre machos e fêmeas de uma espécie.
Um exemplo: a resistência aos antibióticos
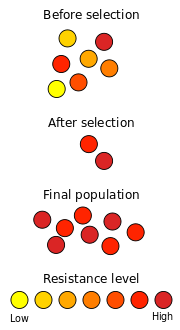
Um exemplo bem conhecido de selecção natural em acção é o desenvolvimento de resistência a antibióticos em microrganismos. Desde a descoberta de penicilina em 1928 por Alexander Fleming , os antibióticos têm sido usados para combater bactérias doenças. Populações naturais de bactérias contêm, entre os seus grandes números de membros individuais, uma variação considerável em seu material genético, principalmente como resultado de mutações. Quando exposto a antibióticos, a maioria das bactérias morrem rapidamente, mas alguns podem ter mutações que os tornam um pouco menos susceptíveis. Se a exposição aos antibióticos é curto, esses indivíduos vão sobreviver ao tratamento. Esta eliminação seletiva de indivíduos mal adaptados a partir de uma população é a seleção natural.
Estas bactérias sobreviventes vão depois reproduzir novamente, produzindo a próxima geração. Devido à eliminação dos indivíduos mal adaptados na geração passado, esta população contém mais bactérias que têm alguma resistência contra o antibiótico. Ao mesmo tempo, novas mutações ocorrem, contribuindo nova variação genética para a variação genética existente. As mutações espontâneas são muito raros, e as mutações vantajosas são ainda mais raros. No entanto, as populações de bactérias são grandes o suficiente para que alguns indivíduos terão mutações benéficas. Se uma nova mutação reduz a sua susceptibilidade a um antibiótico, estes indivíduos têm maior probabilidade de sobreviver quando próxima confrontado com este antibiótico. Dado o tempo suficiente, e a exposição repetida ao antibiótico, uma população de bactérias resistentes aos antibióticos irão emergir.
O uso generalizado e uso indevido de antibióticos resultou em aumento da resistência microbiana aos antibióticos em uso clínico, a tal ponto que o Staphylococcus aureus resistente à meticilina (MRSA) tem sido descrito como um " superbactéria 'por causa da ameaça que representa para a saúde e sua invulnerabilidade em relação aos medicamentos existentes. Estratégias de resposta incluem tipicamente o uso de antibióticos diferentes, mais fortes; no entanto, a nova estirpes de MRSA têm surgido recentemente que são resistentes mesmo a estes fármacos. Este é um exemplo do que é conhecido como um corrida armamentista evolucionária, na qual as bactérias continuam a desenvolver variedades que são menos suscetíveis aos antibióticos, enquanto que os pesquisadores médicos continuar a desenvolver novos antibióticos que podem matá-los. Uma situação similar ocorre com resistência aos pesticidas nas plantas e insetos. Corridas armamentistas não são necessariamente induzidas pelo homem; Um exemplo bem documentado envolve a elaboração do RNA interferência na via de plantas como meio de imunidade inata contra vírus .
Teoria genética da seleção natural
A seleção natural, por si só é um conceito simples, em que as diferenças de aptidão entre fenótipos desempenhar um papel crucial. É a união da seleção natural como um mecanismo com o material genético como um substrato que oferece mais de poder explicativo da teoria
Direcionalidade da seleção
Quando algum componente de um traço é hereditário, a seleção irá alterar as frequências dos diferentes alelos ou variantes do gene que produz as variantes da característica. A selecção pode ser divididos em três classes, com base no seu efeito sobre a frequência de alelos.
Selecção direccional ocorre quando um determinado alelo tem uma aptidão maior do que outros, o que resulta em um aumento na frequência desse alelo. Este processo pode continuar até que o alelo está fixo e toda a população partes do fenótipo montador. É selecção direccional que é ilustrado no exemplo resistência a antibióticos acima .
Muito mais comum é seleção estabilizadora (também conhecido como selecção purificação), o que reduz a frequência de alelos que têm um efeito prejudicial sobre o fenótipo - isto é, produzir organismos de menor aptidão. Este processo pode continuar até que o alelo é eliminado da população. Resultados da seleção purificadora em recursos genéticos funcionais, tais como genes codificadores de proteínas ou sequências regulatórias, sendo conservado ao longo do tempo devido à pressão seletiva contra as variantes deletérias.
Finalmente, um número de formas de equilibrando selecção existir, que não resultam em fixação, mas manter um alelo a frequências intermédias numa população. Isto pode ocorrer nas espécies diplóides (isto é, aqueles que têm dois pares de cromossomas) quando indivíduos heterozigotos, que têm diferentes alelos em cada cromossoma em um único locus genético, possuem uma aptidão superior a indivíduos homozigotos que têm dois dos mesmos alelos. Isto é chamado vantagem heterozigoto ou sobredominância, dos quais o exemplo mais conhecido é o da malária resistência observada em humanos heterozigotos que carregam apenas uma cópia do gene para a anemia falciforme . Manutenção de variação alélica pode também ocorrer através selecção disruptiva ou diversificando, o que favorece genótipos que partem da média em qualquer direcção (isto é, o oposto do sobredominância), e pode resultar numa distribuição bimodal de valores traço. Finalmente, equilibrando selecção pode ocorrer através dependente da frequência de selecção, em que a aptidão de um fenótipo particular depende da distribuição de outros fenótipos na população. Os princípios da teoria dos jogos têm sido aplicados para entender as distribuições de fitness nestas situações, particularmente no estudo da seleção de parentesco e da evolução do altruísmo recíproco.
Seleção e variação genética
Uma porção de todos variação genética é funcionalmente neutra em que não produz qualquer efeito fenotípico ou diferença significativa na aptidão; a hipótese de que esta variação representa uma grande fração do observado diversidade genética é conhecida como a teoria neutra da evolução molecular e foi originado por Motoo Kimura. Variação neutra já foi pensado para abranger a maior parte da variação genética em ADN não codificante, que foi levantada a hipótese de ser composto por " DNA lixo ". No entanto, mais recentemente, o papel funcional de ADN não codificante, tais como as funções de regulação e de desenvolvimento de RNA produtos de genes, tem sido estudada em profundidade; grandes partes de sequências de ADN não codificantes de proteínas são altamente conservadas sob uma forte selecção de purificação e, portanto, não variam muito de indivíduo para indivíduo, o que indica que as mutações nestas regiões têm consequências deletérias. Quando a variação genética não resulta em diferenças de fitness, a seleção não pode afetar diretamente a freqüência de tal variação. Como resultado, a variação genética nesses locais será maior do que em locais onde a variação faz influência de fitness.
Equilíbrio seleção mutação
A selecção natural resulta na redução da variação genética através da eliminação de indivíduos mal adaptados e, consequentemente, das mutações que causam a má adaptação. Ao mesmo tempo, novas mutações ocorrem, resultando numa equilíbrio mutação-seleção. O resultado exacto dos dois processos depende tanto da taxa a que ocorrem novas mutações e na força da selecção natural, que é uma função de como a mutação desfavorável revela-se. Por conseguinte, as alterações na taxa de mutação ou a pressão de selecção irá resultar num saldo mutação-selecção diferente.
Ligação genética
Ligação genética ocorre quando o loci de dois alelos estão ligados, ou em estreita proximidade uns dos outros no cromossoma. Durante a formação de gâmetas, recombinação dos resultados materiais genéticos na reorganização dos alelos. No entanto, a possibilidade de que uma tal remodelação ocorre entre dois alelos depende da distância entre esses alelos; Quanto mais próximos os alelos estão um para o outro, o que é menos provável que um tal remodelação irá ocorrer. Por conseguinte, quando a selecção do alvo, um alelo, isto resulta automaticamente na selecção do outro alelo bem; através deste mecanismo, a selecção pode ter uma forte influência sobre os padrões de variação no genoma.
Varreduras selectivos ocorrer quando um alelo torna-se mais comum na população como um resultado de selecção positiva. Como a prevalência de um aumento de alelos, alelos ligados também pode se tornar mais comum, se eles são neutros ou mesmo ligeiramente deletério. Isto é chamado hitchhiking genética. Um forte resultados de varrimento selectivos em uma região do genoma onde o positivamente seleccionadas haplótipo (alelo e os seus vizinhos) são essencialmente os únicos que existem na população.
Se uma limpeza seletiva ocorreu ou não pode ser investigado medindo desequilíbrio de ligação, ou se um dado haplótipo é sobre-representadas na população. Normalmente, recombinação genética resultar num rearranjo dos diferentes alelos dentro de um haplótipo, e nenhum dos haplótipos irá dominar a população. No entanto, durante uma limpeza seletiva, selecção por um alelo específico irá também resultar na selecção de alelos vizinhos. Portanto, a presença de forte desequilíbrio de ligação pode indicar que houve uma varredura selectiva "recente", e isto pode ser utilizado para identificar locais recentemente sob selecção.
Seleção de fundo é o oposto de uma limpeza seletiva. Se um local específico experimenta selecção purificação forte e persistente, a variação ligada tenderá a ser eliminadas juntamente com isso, a produção de uma região do genoma de baixa variabilidade global. Porque selecção fundo é um resultado de novas mutações deletérias, que podem ocorrer em qualquer aleatoriamente haplótipos, mas não produz nenhuma desequilíbrio de ligação.
Evolução por meio de seleção natural
Um pré-requisito para a seleção natural resultar em evolução adaptativa, novas características e especiação, é a presença de hereditárias variação genética que resulta em diferenças de fitness. A variação genética é o resultado de mutações, recombinações e alterações no cariótipo (o número, forma, tamanho e disposição interna do cromossomos). Qualquer uma dessas alterações pode ter um efeito que é altamente vantajosa ou desvantajosa altamente, mas grandes efeitos são muito raros. No passado, a maioria das alterações no material genético foram considerados neutros ou próximo de neutro, porque eles ocorreram em o DNA não codificante ou resultou numa substituição sinônimo. No entanto, pesquisas recentes sugerem que muitas mutações no DNA não-codificante tem efeitos deletérios ligeiras. Embora ambas as taxas de mutação e efeitos médios de fitness de mutações são dependentes do organismo, as estimativas de dados em seres humanos descobriram que uma maioria das mutações são um pouco prejudicial.

Pela definição de fitness, os indivíduos com maior aptidão são mais propensos a contribuir prole para a próxima geração, enquanto que os indivíduos com menor aptidão são mais propensos a morrer mais cedo ou deixar de reproduzir. Como resultado, os alelos que em resultado médio em maior aptidão tornaram mais abundantes na próxima geração, enquanto os alelos que geralmente reduzir aptidão tornaram mais raros. Se as forças de selecção continuam a ser o mesmo para muitas gerações, os alelos benéficos tornam-se mais e mais abundante, até que eles dominam a população, enquanto os alelos com uma aptidão menor desaparecer. Em cada geração, novas mutações e recombinações surgem espontaneamente, produzindo um novo espectro de fenótipos. Por isso, cada nova geração será enriquecido com a crescente abundância de alelos que contribuem para esses traços que foram favorecidas pela seleção, reforçando essas características ao longo de gerações sucessivas.
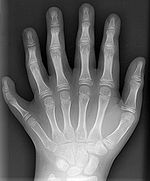
Algumas mutações ocorrem nos chamados genes reguladores. Alterações em que estes podem ter grandes efeitos sobre o fenótipo do indivíduo porque eles regulam a função de muitos outros genes. A maioria, mas não todas, as mutações nos genes reguladores resultar em não-viáveis zigotos. Exemplos de mutações ocorrem em reguladoras não letais Genes HOX em seres humanos, o que pode resultar numa costela cervical ou polydactyly, um aumento no número de dedos ou dedos do pé. Quando essas mutações resultam em uma aptidão maior, a seleção natural favorecerá esses fenótipos ea nova característica vai se espalhar na população.
Traços estabelecidos não são imutáveis; traços que têm alta aptidão em um contexto ambiental pode ser muito menos ajuste, se as condições ambientais mudarem. Na ausência de selecção natural para preservar uma tal característica, torna-se mais variável e deteriorar-se ao longo do tempo, resultando possivelmente numa vestigial manifestação do traço. Em muitas circunstâncias, a estrutura aparentemente vestigial pode reter uma funcionalidade limitada, ou podem ser co-optado para outras características vantajosas, em um fenómeno conhecido como pré-adaptação. Um exemplo famoso de uma estrutura vestigial, o olho do cego rato-toupeira, acredita-se reter a função em percepção fotoperíodo.
Especiação
Especiação requer acasalamento seletivo, que resultam em uma reduzida fluxo gênico. Acoplamento selectivo pode ser o resultado de, por exemplo, uma alteração no ambiente físico (isolamento físico por uma barreira extrínseca), quer por selecção sexual, resultando em acasalamento. Com o tempo, estes subgrupos pode divergir radicalmente a tornar-se diferentes espécies, seja por causa de diferenças de pressões seletivas sobre os diferentes subgrupos, ou porque diferentes mutações surgem espontaneamente nas diferentes populações, ou por causa de efeito fundador - alguns alelos potencialmente benéficos podem, por acaso, estar presentes em apenas um ou outro dos dois subgrupos quando primeiro se separar. Um mecanismo menos conhecido da especiação ocorre via hibridização, bem documentada em plantas e ocasionalmente observada em grupos ricos em espécies de animais, tais como peixes ciclídeos. Tais mecanismos de especiação rápida pode refletir um mecanismo de mudança evolutiva conhecida como equilíbrio pontuado , o que sugere que a mudança evolutiva e particularmente especiação tipicamente acontece rapidamente após a interrupção longos períodos de imobilização.
As alterações genéticas dentro de grupos resultar no aumento da incompatibilidade entre os genomas dos dois subgrupos, reduzindo assim o fluxo de genes entre os grupos. O fluxo gênico deixem efectivamente quando as mutações distintas que caracterizam cada subgrupo se tornar fixo. Como apenas duas mutações podem resultar na formação de espécies: se cada mutação tem um efeito neutro ou positivo sobre a aptidão quando ocorrem separadamente, mas um efeito negativo quando elas ocorrem em conjunto, então a fixação desses genes nos respectivos subgrupos levará a dois reprodutivamente isolado populações. De acordo com o conceito de espécie biológica, estas serão duas espécies diferentes.
Desenvolvimento histórico

Teorias pré-darwinianas
Vários filósofos antigos expressa a idéia de que natureza produz uma enorme variedade de criaturas, aparentemente de forma aleatória, e que apenas aquelas criaturas que sobrevivem gerir a fornecer para si e para reproduzir com sucesso; Exemplos bem conhecidos incluem Empédocles e seu sucessor intelectual, Lucrécio, enquanto idéias relacionadas foram posteriormente refinado por Aristóteles . A luta pela existência foi mais tarde descrita por al-Jahiz no século 9. Tais argumentos clássicos foram reintroduzidos no século 18 por Pierre Louis Maupertuis e outros, incluindo Charles Darwin avô Erasmus Darwin. Embora estes precursores teve uma influência sobre Darwinismo, que mais tarde teve pouca influência sobre a trajetória do pensamento evolutivo após Charles Darwin.
Até o início do século 19, o vista nas sociedades ocidentais predominante era que as diferenças entre indivíduos de uma espécie foram partidas desinteressantes de sua Ideal platônico (ou typus) de espécies criadas. No entanto, a teoria de uniformitarianism em geologia promoveu a idéia de que as forças simples, fracos poderia agir de forma contínua durante longos períodos de tempo para produzir mudanças radicais na paisagem da Terra. O sucesso desta teoria aumentou a consciência sobre a grande escala de tempo geológico e torna plausível a idéia de que pequenas mudanças, praticamente imperceptíveis em sucessivas gerações poderia produzir consequências na escala de diferenças entre as espécies. Início do século 19 evolucionistas como Jean Baptiste Lamarck sugeriu a herança de características adquiridas como um mecanismo de mudança evolutiva; características adaptativas adquiridas por um organismo durante a sua vida poderia ser herdado por descendência desse organismo, eventualmente causando transmutação das espécies. Esta teoria tem vindo a ser conhecido como Lamarckismo e era uma influência sobre as ideias anti-genéticas do Stalinista soviético biólogo Trofim Lysenko.
A hipótese de Darwin
Entre 1842 e 1844, Charles Darwin esboçou sua teoria da evolução pela seleção natural como uma explicação para adaptação e especiação. Ele definiu a seleção natural como o "princípio por que cada leve variação [de um traço], se útil, é preservada". O conceito era simples, mas poderosa: os indivíduos melhor adaptados aos seus ambientes são mais propensos a sobreviver e se reproduzir. Contanto que há alguma variação entre eles, haverá uma selecção inevitável de indivíduos com as variações mais vantajosas. Se as variações são herdadas, em seguida, o sucesso reprodutivo diferencial vai levar a uma evolução progressiva de determinadas populações de uma espécie, e as populações que evoluem para ser suficientemente diferente pode eventualmente tornar-se espécies diferentes.
As idéias de Darwin foram inspirados pelas observações que fizera em A viagem do Beagle, e pelo trabalho de dois economistas. O primeiro foi Thomas Malthus , que em Um Ensaio sobre o Princípio da População, observou que a população (se não for controlada) aumenta exponencialmente ao passo que a oferta de alimentos cresce apenas aritmeticamente; portanto, inevitáveis limitações de recursos teria implicações demográficas, levando a uma "luta pela existência", em que somente os mais aptos sobreviveriam. O segundo foi Adam Smith , que, em A Riqueza das Nações, identificou um mecanismo de regulação nos mercados livres, que ele se referia como a " mão invisível ", o que sugere que os preços auto-ajustar de acordo com suprimentos e demanda. Assim, para Darwin, o desastre que deveria ocorrer de acordo com Malthus foi mantida sob controle e constantemente melhorado por concorrência (ou lei da seleção).
Uma vez que a teoria tinha sido formulada, Darwin era meticuloso sobre coleta e provas de refino, compartilhando suas idéias só com alguns amigos; ele se inspirou para publicar após o naturalista Alfred Russel Wallace concebido independentemente do princípio e descreveu-o em um ensaio que ele enviou para Darwin. Um acordo foi feito (sem o conhecimento de Wallace) para apresentar seu ensaio e dois escritos inéditos curtas de Darwin para o Linnean Society anunciando co-descoberta do princípio em julho 1858; Darwin publicou um relato mais detalhado de suas provas e conclusões em A Origem das Espécies em 1859. Na 6ª edição de A Origem das Espécies de Darwin reconheceu que outros - nomeadamente William Charles Wells em 1813, e Patrick Matthew em 1831 - tinha proposto teorias semelhantes, mas não apresentaram-los totalmente ou em publicações científicas notáveis.
Darwin pensava da seleção natural, por analogia, como os agricultores selecionar culturas ou pecuárias de criação, que ele chamou de seleção artificial; em seus primeiros manuscritos que ele se referia a uma "natureza" que faria a seleção. Na altura, outros mecanismos de evolução tal como evolução por deriva genética ainda não foram explicitamente formulado, e Darwin percebeu que a seleção era provavelmente apenas uma parte da história: ". Estou convencido de que [ele] tem sido os principais, mas não meio exclusivo de modificação" Para Darwin e seus contemporâneos, a seleção natural foi, assim, essencialmente sinônimo de evolução por seleção natural . Depois da publicação de A Origem das Espécies, pessoas educadas geralmente aceite que a evolução tivesse ocorrido em alguma forma. No entanto, a seleção natural permaneceu controversa como um mecanismo, em parte porque ele era considerado muito fraco para explicar a gama de características observadas dos organismos vivos, e em parte porque até mesmo partidários da evolução recusou-se a sua "não dirigida" e de natureza não progressiva, uma response that has been characterized as the single most significant impediment to the idea's acceptance. No entanto, alguns pensadores abraçou entusiasticamente a seleção natural; depois de ler Darwin, Herbert Spencer introduziu o termo sobrevivência do mais apto , que se tornou um resumo popular da teoria. Embora a frase ainda é muito utilizado por não-biólogos, biólogos modernos evitá-lo, porque é tautológica se mais apto é lido como significando funcionalmente superior, e é aplicado a indivíduos, e não considerado como uma quantidade média sobre populações. Em uma carta ao Charles Lyell em 1860 de setembro de Darwin lamenta a utilização do termo "seleção natural", preferindo o termo "Natural Preservação".
Síntese evolutiva moderna
Somente após a integração de uma teoria da evolução com uma apreciação estatística complexa de monge austríaco leis de Gregor Mendel 're-descoberta "de herança que a seleção natural se tornar geralmente aceite pelos cientistas. O trabalho de Ronald Fisher (que desenvolveu a linguagem da matemática e da seleção natural em termos dos processos genéticos subjacentes), JBS Haldane (que introduziu o conceito de "custo" da seleção natural), Sewall Wright (que elucidou a natureza de seleção e adaptação), Theodosius Dobzhansky (que estabeleceu a ideia de que a mutação, criando genética diversidade, forneceu a matéria-prima para a seleção natural), William Hamilton (que concebeu de seleção de parentesco), Ernst Mayr (que reconheceu a importância fundamental de isolamento reprodutivo para a especiação) e muitos outros formaram a síntese evolutiva moderna. Esta síntese cimentou a seleção natural como base da teoria da evolução, onde permanece até hoje.
Impacto da idéia
Darwin 's idéias, juntamente com os de Adam Smith e Karl Marx , teve uma profunda influência sobre o pensamento do século 19. Talvez a afirmação mais radical da teoria da evolução através da seleção natural é que "formas elaboradamente construídas, tão diferentes um do outro, e dependentes um do outro em uma maneira tão complexa" evoluíram de formas mais simples de vida por alguns princípios simples. Esta alegação inspirou alguns dos mais ardorosos defensores de Darwin-e provocou a mais profunda oposição. O radicalismo da seleção natural, de acordo com Stephen Jay Gould , estava ao seu alcance para "destronar alguns dos confortos mais profundos e mais tradicionais do pensamento ocidental". Em concreto, contestou as crenças de longa data em tais conceitos como um lugar especial e exaltado para os seres humanos no mundo natural e um criador benevolente cujas intenções foram refletidas em ordem e de design da natureza.
A teoria social e psicológico
As implicações sociais da teoria da evolução pela seleção natural também se tornou fonte de contínua controvérsia. Friedrich Engels , um alemão filósofo político e co-autor da ideologia do comunismo , escreveu em 1872 que "Darwin não sabia o que uma sátira amarga ele escreveu sobre a humanidade quando ele mostrou que a livre concorrência, a luta pela existência, o que os economistas comemorar como a maior conquista histórica, é o estado normal do reino animal ". Interpretação da seleção natural como necessariamente "progressista", levando ao aumento da 'avança' em inteligência e civilização, foi usada como uma justificativa para o colonialismo e as políticas de eugenia, bem como as posições sócio-políticas mais amplas agora descrito como darwinismo social. Konrad Lorenz ganhou o Nobel Prêmio de Fisiologia ou Medicina em 1973 por sua análise do comportamento animal em termos do papel da seleção natural (em particular a seleção de grupo). No entanto, na Alemanha, em 1940, nos escritos que ele posteriormente rejeitadas, ele usou a teoria como justificativa para as políticas do Nazi estado. Ele não escreveu "... a seleção para resistência, heroísmo e utilidade social ... deve ser efectuada por qualquer instituição humana, se a humanidade, na falta de fatores seletivos, é para ser arruinada por degeneração induzida por domesticação. A idéia racial como a base do nosso Estado já realizou muito a esse respeito. " Outros desenvolveram ideias que as sociedades humanas e da cultura evoluem por meio de mecanismos que são análogas às que se aplicam a evolução das espécies.
Mais recentemente, o trabalho entre antropólogos e psicólogos levou ao desenvolvimento de sociobiologia e mais tarde a psicologia evolucionista, um campo que tenta explicar características de psicologia humana em termos de adaptação ao ambiente ancestral. O exemplo mais proeminente tal, nomeadamente avançada no trabalho inicial de Noam Chomsky e depois por Steven Pinker, é a hipótese de que o cérebro humano está adaptado para adquirir as regras gramaticais de linguagem natural. Outros aspectos do comportamento humano e as estruturas sociais, de normas culturais específicas, tais como evitar o incesto a padrões mais amplos, como os papéis de gênero, foram supor para ter origens similares como adaptações ao ambiente precoce no qual os humanos modernos evoluíram. Por analogia com a ação da seleção natural nos genes, o conceito de memes - "unidades de transmissão cultural", ou equivalentes de cultura de genes submetidos à seleção e recombinação - surgiu, descrita pela primeira vez nesta forma por Richard Dawkins e posteriormente expandida por filósofos tais como Daniel Dennett como explicações para atividades culturais complexas, incluindo humanos consciência. extensões da teoria da seleção natural para uma ampla gama de fenômenos culturais, foram distintamente controverso e não são amplamente aceitos.
A teoria da informação e sistemas
Em 1922, Alfred Lotka propôs que a seleção natural pode ser entendido como um princípio físico que poderia ser energicamente quantificado, um conceito que foi desenvolvido mais tarde por Howard Odum como o princípio segundo o qual a potência máxima sistemas evolutivos com vantagem selectiva maximizar a taxa de transformação de energia útil. Tais conceitos são, por vezes, relevante no estudo da aplicados termodinâmica .
Os princípios da seleção natural ter inspirado uma variedade de técnicas computacionais, tais como "soft" vida artificial, que simulam processos seletivos e podem ser altamente eficiente em "Adaptação" entidades para um ambiente definido por uma especificada função de fitness. Por exemplo, uma classe de heurísticas de otimização de algoritmos conhecidos como algoritmos genéticos, desenvolvidas por John Holland em 1970 e expandida por David Goldberg, identificar as melhores soluções por reprodução simulada e mutação de uma população de soluções definidas por uma inicial distribuição de probabilidade . Tais algoritmos são particularmente úteis quando aplicados a problemas cuja paisagem solução é muito áspera ou tem muitos mínimos locais.