

Chromatophore
Sabías ...
Esta selección se hace para las escuelas por caridad para niños leer más . Apadrinar un niño para hacer una diferencia real.

Cromatóforos son que reflejan la luz que contiene pigmento y células que se encuentran en anfibios, peces , reptiles , crustáceos , y cefalópodos. Ellos son en gran parte responsables de la generación de la piel y color de ojos en sangre fría animales y se generan en el cresta neural durante el desarrollo embrionario. Cromatóforos maduros se agrupan en subclases basadas en su color (más correctamente " hue ") con luz blanca: xanthophores (amarillo), eritróforos (rojo), iridophores ( reflexivo / iridiscente), leucoforos (blanco), melanóforos (negro / marrón) y cyanophores (azul). El término también puede referirse a vesículas de color, asociadas a la membrana que se encuentran en algunas formas de bacterias fotosintéticas.
Algunas especies pueden cambiar rápidamente de color a través de mecanismos que translocate pigmento y reorientar placas reflectantes dentro cromatóforos. Este proceso, a menudo utilizado como un tipo de camuflaje, se llama cambio de color fisiológica. Los cefalópodos como pulpo tienen órganos cromatóforos complejos controlados por los músculos para lograrlo, mientras que los vertebrados tales como camaleones generan un efecto similar por la señalización celular. Tales señales pueden ser hormonas o neurotransmisores y pueden ser iniciadas por los cambios en el estado de ánimo, la temperatura, el estrés o cambios visibles en el entorno local.
A diferencia de los animales de sangre fría, los mamíferos y las aves tienen una sola clase de los cromatóforos como tipo de células: la melanocitos. El equivalente a sangre fría, melanóforos, son estudiados por los científicos para comprender las enfermedades humanas y se utiliza como una herramienta de descubrimiento de fármacos.
Clasificación
Invertebrados células pigmentarias que devengan fueron descritos por primera vez como chromoforo en una revista científica italiana en 1819. El término fue adoptado chromatophore más tarde como el nombre de células de pigmento que lleva derivados de la cresta neural de sangre fría vertebrados y cefalópodos. La palabra en sí proviene del griego palabras KHROMA (χρωμα) que significa "color", y phoros (φορος) que significa "llevar". Por el contrario, se adoptó la palabra chromatocyte (cyte o κυτε ser griego para "célula") para las células responsables de color que se encuentra en las aves y mamíferos. Sólo uno de estos tipo de célula, el melanocito, se ha identificado en estos animales.
No fue sino hasta la década de 1960 que la estructura y coloración de cromatóforos se entienden suficientemente bien como para permitir el desarrollo de un sistema de sub-clasificación basada en su apariencia. Este sistema de clasificación persiste hasta nuestros días a pesar de que los estudios más recientes han revelado que ciertos bioquímicos aspectos de los pigmentos pueden ser más útiles para una comprensión científica de cómo funcionan las células.
Color-producción cae en clases distintas: biochromes, schemochromes . Los biochromes incluyen pigmentos verdaderos, como carotenoides y pteridinas. Estos pigmentos absorben selectivamente partes de la espectro de luz visible que hace que la luz blanca al tiempo que permite otra longitudes de onda para llegar al ojo del observador. Schemochromes, también conocidos como "colores estructurales", producen coloración al reflejar algunas longitudes de onda (colores) de luz y la transmisión de otros, haciendo que las ondas de luz para interferir dentro de la estructura o por la dispersión de luz que cae sobre ellos.
Mientras que todos los cromatóforos contienen pigmentos o estructuras reflectantes (excepto cuando se ha producido un genética mutación que resulta en un trastorno como el albinismo ), no todas las células de pigmento que contiene son cromatóforos. Hemo, por ejemplo, es un Biochrome responsable de la aparición de color rojo de la sangre. Se encuentra principalmente en glóbulos rojos (eritrocitos), que se generan en la médula ósea durante toda la vida de un organismo, en lugar de ser formada durante el desarrollo embriológico. Por lo tanto los eritrocitos no se clasifican como cromatóforos.

Xanthophores y eritróforos
Cromatóforos que contienen grandes cantidades de pigmentos amarillos de pteridina se nombran xanthophores y aquellos con un exceso de rojo / carotenoides naranja denominan eritróforos. Se descubrió que pteridina y que contiene carotenoide vesículas se encuentran a veces dentro de la misma célula, y que el color general depende de la relación de los pigmentos rojos y amarillos. Por lo tanto la distinción entre estos tipos cromatóforos es esencialmente arbitrario. La capacidad de generar pteridinas desde trifosfato de guanosina es una característica común a la mayoría de cromatóforos, pero xanthophores parecen tener rutas bioquímicas suplementarios que resultan en una acumulación excesiva de pigmento amarillo. En contraste, los carotenoides son metabolizados de la dieta y transportados a eritróforos. Esto fue demostrado por primera vez por la cría de ranas verdes normalmente en una dieta de restringido-caroteno grillos. La ausencia de caroteno en la dieta de la rana significó el rojo / naranja carotenoide de color "filtro" no estaba presente en eritróforos. Esto resultó en la rana que aparece azul en el color, en lugar de verde.
Iridophores y leucoforos
Iridophores, a veces también llamados guanophores, son células de pigmento que reflejan la luz utilizando placas de chemochromes cristalinos hechos de guanina. Cuando se ilumina generan colores iridiscentes, debido a la difracción de la luz dentro de las placas apiladas. Orientación de la schemochrome determina la naturaleza del color observado. Mediante el uso de biochromes como filtros de colores, iridophores crean un efecto óptico conocido como Tyndall o La dispersión de Rayleigh, brillante producir azul o colores verdes.
Un tipo relacionado de chromatophore, la leucophore, se encuentra en algunos peces, particularmente en el tapetum lucidum. Como iridophores, utilizan cristalina purinas (a menudo guanina) para reflejar la luz. A diferencia iridophores, sin embargo, leucoforos tienen cristales más organizados que reducen la difracción. Dada una fuente de luz blanca, producen una brillo blanco. Al igual que con xanthophores y eritróforos, en el pescado la distinción entre iridophores y leucoforos no siempre es obvia, pero en general iridophores se consideran para generar iridiscentes o metálicos colores mientras leucoforos producen tonos blancos reflectantes.
Melanóforos
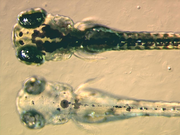
Melanóforos contienen eumelanina, un tipo de melanina, que aparece negro u oscuro marrón debido a sus cualidades de absorción de luz. Se empaqueta en vesículas llamados melanosomas y distribuidos por toda la célula. La eumelanina se genera a partir tirosina en una serie de reacciones químicas catalizadas. Es una sustancia química compleja que contiene unidades de dihidroxiindol-2- dihidroxiindol y el ácido carboxílico con un poco de anillos de pirrol. La enzima clave en la síntesis de melanina es tirosinasa. Cuando esta proteína es defectuoso, no se puede generar melanina resultante en ciertos tipos de albinismo. En algunas especies de anfibios que hay otros pigmentos empaquetados junto eumelanina. Por ejemplo, una novela profunda pigmento de color rojo fue identificado en los melanóforos de phyllomedusine ranas . Este fue identificado posteriormente como pterorhodin, una pteridina dímero que se acumula alrededor de eumelanina. Si bien es probable que otras especies menos estudiadas tienen pigmentos melanophore complejos, no es menos cierto que la mayoría de melanóforos estudiados hasta la fecha no contener exclusivamente la eumelanina.
Los seres humanos tienen sólo una clase de células de pigmento, el equivalente de los mamíferos de melanóforos, para generar la piel, el cabello y color de ojos. Por esta razón, y debido a la gran cantidad y color de contraste de las células por lo general hacen muy fácil de visualizar, melanóforos son con mucho el chromatophore más ampliamente estudiado. Sin embargo, hay diferencias entre la biología de melanóforos y melanocitos. Además de eumelanina, los melanocitos pueden generar un pigmento amarillo / rojo llamado feomelanina.

Cyanophores
En 1995 se demostró que los vibrantes colores azules en algunos tipos de El pez mandarín no son generadas por schemochromes. En cambio, una Biochrome cian de la naturaleza química desconocida es responsable. Este pigmento, que se encuentra dentro de las vesículas en al menos dos especies de peces callionymid, es muy inusual en el animal de reino, como todos los demás colorantes azules hasta ahora investigados son schemochromatic. Por lo tanto un tipo chromatophore novela, el cyanophore, se propuso. Aunque parecen inusual en su restricción taxonómica, puede haber cyanophores (así como los tipos más inusuales cromatóforos) en otros peces y anfibios. Por ejemplo, cromatóforos de colores brillantes con pigmentos indefinidos se han observado en tanto ranas venenosas y ranas de cristal.
Translocación Pigmento
Muchas especies tienen la capacidad de trasladar el pigmento dentro de cromatóforos, resultando en un cambio aparente en color. Este proceso, conocido como cambio de color fisiológica, es el más ampliamente estudiado en melanóforos, ya que la melanina es el pigmento más oscuro y más visible. En la mayoría de especies con una relativamente delgada dermis, los melanóforos dérmicos tienden a ser plana y cubrir una gran superficie. Sin embargo, en los animales con capas dérmicas de espesor, tales como reptiles adultos, melanóforos dérmicos a menudo forman unidades tridimensionales con otros cromatóforos. Estas unidades cromatóforos dérmica (DCU) se componen de un xanthophore superior o erythrophore capa, a continuación, una capa iridophore, y finalmente una capa de melanóforos de cesta con los procesos que cubren las iridophores.
Ambos tipos de melanóforos dérmicos son importantes en el cambio de color fisiológico. Melanóforos dérmicos planos a menudo superponer otros cromatóforos así que cuando el pigmento se dispersa por toda la célula de la piel aparece oscuro. Cuando el pigmento se agrega hacia el centro de la célula, los pigmentos en otros cromatóforos están expuestos a la luz y la piel adquiere su tonalidad. Del mismo modo, después de la agregación de melanina en DCU, la piel aparece verde a través xanthophore (amarillo) de filtrado de la luz dispersada desde la capa iridophore. En la dispersión de la melanina, la luz ya no es dispersa y la piel aparece oscuro. Como los otros chomatophores biocromática también son capaces de translocación pigmento, los animales con múltiples tipos cromatóforos pueden generar una espectacular variedad de colores de piel, haciendo un buen uso de los efectos de la división.,

El control y la mecánica de la rápida translocación el pigmento se ha estudiado bien en un número de diferentes especies, en particular anfibios y pez teleósteo., se ha demostrado que el proceso puede estar bajo hormonal , control neuronal o ambos. Neuroquímicos que se sabe que trasladar pigmento incluyen noradrenalina, a través de su receptor en la superficie en melanóforos. Las principales hormonas involucradas en la regulación de la translocación parece ser el melanocortinas, melatonina y hormona concentradora de melanina (MCH), que se producen principalmente en la hipófisis, glándula pineal y el hipotálamo respectivamente. Estas hormonas también pueden ser generados en una paracrina por las células en la piel. En la superficie de la melanophore se ha demostrado que las hormonas para activar específica G-receptores acoplados a proteínas que, a su vez, transducen la señal en la célula. Melanocortinas resultan en la dispersión de pigmento, mientras que la melatonina y los resultados de SMI en la agregación.
Numerosos receptores de melanocortina, SMI y la melatonina se han identificado en peces y ranas, incluyendo una homólogo de MC1R, un receptor de melanocortina conocido para regular piel y color de cabello en los seres humanos. Dentro de la célula, monofosfato de adenosina cíclico (cAMP) ha demostrado ser un importante segundo mensajero de la translocación pigmento. A través de un mecanismo aún no se entiende completamente, las influencias de AMPc otras proteínas tales como la proteína quinasa A para conducir motores moleculares que llevan pigmento que contiene vesículas a lo largo de ambos microtúbulos y microfilamentos.,,
Adaptación de fondo
La mayoría de los peces, los reptiles y anfibios se someten a un cambio de color fisiológica limitada en respuesta a un cambio en el medio ambiente. Este tipo de camuflaje, conocido como adaptación de fondo, por lo general aparece como un leve oscurecimiento o decoloración de tono de la piel de aproximadamente imitar el tono del entorno inmediato. Se ha demostrado que el proceso de adaptación de fondo es dependiente de la visión (parece que el animal necesita para ser capaz de ver el entorno para adaptarse a ella), y que la translocación de melanina en melanóforos es el factor principal en el cambio de color. Algunos animales, como los camaleones y anoles, tienen una respuesta de adaptación fondo altamente desarrollada capaz de generar un número de diferentes colores muy rápidamente. Se han adaptado la capacidad de cambiar de color en respuesta a la temperatura, estado de ánimo, los niveles de estrés y señales sociales, en lugar de simplemente imitar su entorno.

Desarrollo
Durante los vertebrados el desarrollo embrionario, cromatóforos son uno de un número de tipos de células generada en el cresta neural, una tira emparejado de las células que surgen en los márgenes de la tubo neural. Estas células tienen la capacidad de migrar largas distancias, lo que permite cromatóforos para poblar muchos órganos del cuerpo, incluyendo la piel, los ojos, el oído y el cerebro. Al salir de la cresta neural en oleadas, cromatóforos toman ya sea una ruta dorsolateral través de la dermis, que entra en el ectodermo a través de pequeños orificios en la lámina basal, o una ruta entre el ventromedial somitas y el tubo neural. La excepción a esto es los melanóforos del epitelio pigmentario de la retina del ojo. Estos no se derivan de la cresta neural, en lugar de una evaginación del tubo neural genera la copa óptica que, a su vez, forma la retina .
¿Cuándo y cómo células precursoras chromatophore multipotentes (llamados chromatoblasts) desarrollan en sus subtipos hija es un área de investigación en curso. Es conocido en embriones de pez cebra, por ejemplo, que por 3 días después de fertilización cada una de las clases de células que se encuentran en los peces adultos - melanóforos, xanthophores y iridophores - ya están presentes. Los estudios que utilizan peces mutantes han demostrado que factores de transcripción como kit, SOX10 y MITF son importantes en el control de la diferenciación chromatophore. Si estas proteínas son defectuosos, cromatóforos pueden ser regional o totalmente ausente, resultando en una trastorno leucistic.
Aplicaciones prácticas
Además de la investigación básica en una mejor comprensión de los propios cromatóforos, las células se utilizan con fines de investigación aplicada. Por ejemplo, las larvas de pez cebra se utilizan para estudiar cómo cromatóforos organizan y se comunican para generar con precisión el patrón de rayas horizontales regulares como se ve en peces adultos. Esto se ve como una útil sistema modelo para entender los patrones en el evolutiva campo de la biología del desarrollo. Biología chromatophore también se ha utilizado para modelar afección o enfermedad humana, incluyendo el melanoma y el albinismo. Recientemente, el gen responsable de la cepa de pez cebra de oro-melanophore específica, SLC24A5, fue demostrado que tiene un equivalente humano que se correlaciona fuertemente con color de piel.
Cromatóforos también se utilizan como una biomarcador de ceguera en las especies de sangre fría, como los animales con ciertos defectos visuales no logran fondo adaptarse a ambientes de luz. Homólogos humanos de los receptores que median la translocación pigmento en melanóforos se cree que participar en procesos tales como la supresión del apetito y bronceado, haciéndolos blanco atractivo para fármacos. Por lo tanto, las compañías farmacéuticas han desarrollado un ensayo biológico para identificar rápidamente posibles compuestos bioactivos utilizando melanóforos de la rana de uñas africana . Otros científicos han desarrollado técnicas para el uso de melanóforos como biosensores, y para la detección rápida de la enfermedad (basado en el descubrimiento de que toxina bloquea la tos ferina de agregación de pigmento en melanóforos de pescado). Potencial Se han propuesto aplicaciones militares de cromatóforos mediada por los cambios de color, sobre todo como un tipo de camuflaje activo.
Cromatóforos cefalópodos

Cefalópodos coleoideos tienen complejos "órganos" multicelulares que utilizan para cambiar de color rápidamente. Esto es más notable en colores brillantes calamar, sepias y pulpos. Cada unidad chromatophore se compone de una sola célula chromatophore y numerosos músculos, los nervios, gliales y células de la vaina. Dentro de la célula chromatophore, gránulos de pigmento están encerrados en un saco elástico, llamado el sacculus cytoelastic. Para cambiar el color del animal distorsiona la forma o el tamaño sacculus por la contracción muscular, cambiando su translucidez, la reflectividad o opacidad. Esto difiere de la utilizada en el mecanismo de peces, anfibios y reptiles, en que la forma de la sáculo se está cambiando en vez de una translocación de vesículas de pigmento dentro de la célula. Sin embargo se consigue un efecto similar.
Los pulpos operan en cromatóforos, pantallas cromáticas en forma de onda complejas, lo que resulta en una variedad de combinaciones de colores que cambian rápidamente. Los nervios que operan los cromatóforos se cree que están posicionados en el cerebro, en un orden similar a los cromatóforos cada uno de ellos control. Esto significa que el patrón de cambio de color coincide con el patrón de activación neuronal . Esto puede explicar por qué, como las neuronas se activan uno tras otro, el cambio de color se produce en ondas. Al igual que los camaleones, cefalópodos usan cambio de color fisiológica para la interacción social. También se encuentran entre los más hábiles a la adaptación de fondo, que tiene la capacidad para que coincida con el color y la textura de su medio ambiente local con notable precisión.
Bacterias
Cromatóforos también se encuentran en las membranas de bacterias fototróficas. Se utiliza principalmente para la fotosíntesis, que contienen pigmentos carotenoides y bacterioclorofila. En bacterias púrpuras, tales como Rhodospirillum rubrum las proteínas captadores de luz son intrínsecos a las membranas cromatóforos. Sin embargo, en bacterias verdes del azufre que se disponen en establecimientos especializados complejos de antena llamados clorosomas.