

El potencial de acción
Acerca de este escuelas selección Wikipedia
Esta selección Wikipedia está disponible sin conexión de SOS Children para su distribución en el mundo en desarrollo. Antes de decidir sobre el patrocinio de un niño, ¿por qué no aprender sobre diferentes obras de caridad de patrocinio primero ?
En fisiología, un potencial de acción es un evento de corta duración en el que la eléctrica potencial de membrana de una célula se eleva rápidamente y cae, siguiendo una trayectoria consistente. Los potenciales de acción se producen en varios tipos de células animales , llamados células excitables, que incluyen neuronas, las células musculares, y células endocrinas, así como en algunas células de la planta . En las neuronas, que desempeñan un papel central en la comunicación de célula a célula. En otros tipos de células, su principal función es activar procesos intracelulares. En las células musculares, por ejemplo, un potencial de acción es el primer paso en la cadena de eventos que conducen a la contracción. En células beta del páncreas, que provocan la liberación de insulina . Los potenciales de acción en las neuronas también son conocidos como "los impulsos nerviosos" o "picos" y la secuencia temporal de los potenciales de acción generados por una neurona se llama su "tren de pico". Una neurona que emite un potencial de acción se dice a menudo para "fuego".
Los potenciales de acción son generados por un tipo especial de canales iónicos dependientes de voltaje integrados en una celda de membrana de plasma. Estos canales se cierran cuando el potencial de membrana se encuentra cerca del potencial de reposo de la célula, pero comenzará rápidamente a abrir si el potencial de membrana aumenta hasta un valor umbral definido con precisión. Cuando los canales abiertos, que permiten un flujo hacia dentro de sodio iones, que cambia el gradiente electroquímico, que a su vez produce un nuevo aumento en el potencial de membrana. Esto provoca más canales para abrir, produciendo una mayor corriente eléctrica, y así sucesivamente. El proceso continúa de forma explosiva hasta que todos los canales de iones disponibles están abiertas, lo que resulta en una gran fase de expansión en el potencial de membrana. La rápida afluencia de iones de sodio hace que la polaridad de la membrana plasmática de revertir, y los canales de iones a continuación, inactivan rápidamente. Como los canales de sodio de cerca, los iones de sodio ya no pueden entrar en la neurona, y se transportan activamente fuera de la membrana plasmática. de potasio canales se activan, y hay una corriente hacia el exterior de iones de potasio, devolviendo el gradiente electroquímico para el estado de reposo . Después de que ha ocurrido un potencial de acción, hay un cambio negativo transitorio, llamado afterhyperpolarization o período refractario, debido a las corrientes de potasio adicionales. Este es el mecanismo que evita que un potencial de acción de viajar de nuevo la forma en que acaba de llegar.
En las células animales, hay dos tipos principales de los potenciales de acción, uno de tipo generados por los canales de sodio dependientes de voltaje, el otro de voltaje calcio canales. Potenciales de acción basado en sodio por lo general duran menos de un milisegundo, mientras que los potenciales de acción basados en calcio pueden durar 100 milisegundos o más. En algunos tipos de neuronas, los picos de calcio lentos proporcionan la fuerza motriz para una larga ráfaga de espigas de sodio emitidas rápidamente. En las células del músculo cardíaco, por otro lado, un pico de sodio rápida inicial proporciona una "imprimación" para provocar la rápida aparición de un pico de calcio, que entonces produce la contracción muscular.
Visión de conjunto
Función
Casi todas las células de animales, plantas y hongos función como baterías, en el sentido de que mantienen una diferencia de tensión entre el interior y el exterior de la célula, siendo el interior del polo negativo de la batería. El voltaje de una célula se mide generalmente en milivoltios (mV), o milésimas de voltios. Una tensión típica de una célula animal es -65 mV aproximadamente un quinceavo de un voltio. Dado que las células son tan pequeñas, tensiones de esta magnitud dan lugar a muy fuertes fuerzas eléctricas dentro de la membrana celular.
En la mayoría de las células, las cambios de voltaje muy poco con el tiempo. Hay algunos tipos de células, sin embargo, que son eléctricamente activo en el sentido de que sus voltajes fluctúan. En algunas de ellas, las tensiones a veces muestran muy rápidas fluctuaciones hacia arriba y hacia abajo que tienen una forma estereotipada: estos ciclos para arriba-y-abajo son conocidos como potenciales de acción. Las duraciones de los potenciales de acción varían en una amplia gama, y por consiguiente son señales analógicas. En las células cerebrales de los animales, la totalidad y hacia abajo hasta ciclo pueden tener lugar en menos de una milésima de segundo. En otros tipos de células, el ciclo puede durar varios segundos.
Las propiedades eléctricas de una célula animal se determinan por la estructura de la membrana que lo rodea. La membrana de la célula consiste en una capa de lípidos con moléculas de moléculas de proteína más grandes incrustados en ella. La capa lipídica es altamente resistente a movimiento de los iones cargados eléctricamente, por lo que funciona principalmente como un aislante. Las grandes moléculas de la membrana embebido, en cambio, proporcionan canales a través de los cuales los iones pueden pasar a través de la membrana, y algunas de las moléculas grandes son capaces de moverse activamente tipos específicos de iones de un lado de la membrana al otro.
Proceso en una neurona típica

Todas las células en los tejidos corporales de animales son eléctricamente polarizado - en otras palabras, mantener una diferencia de voltaje a través de la celda de membrana de plasma, conocido como el potencial de membrana. Esta polarización eléctrica resulta de una compleja interacción entre las estructuras de proteínas incrustadas en la membrana llamada bombas de iones y canales iónicos. En las neuronas, los tipos de canales iónicos en la membrana por lo general varía en los diferentes partes de la célula, dando la dendritas, axón, y cuerpo de células diferentes propiedades eléctricas. Como resultado, algunas partes de la membrana de una neurona pueden ser excitable (capaz de generar potenciales de acción), mientras que otros no lo son. La parte más excitable de una neurona es generalmente el axón colina (el punto donde el axón sale del cuerpo de la célula), pero el axón y célula del cuerpo también son excitable en la mayoría de los casos.
Cada parche excitable de la membrana tiene dos importantes niveles de potencial de membrana: la potencial, que es el valor del potencial de membrana mantiene mientras nada perturba la célula, y un valor más alto llamado descansando potencial umbral. En el axón de una neurona típica, el potencial de reposo es de alrededor de -70 milivoltios (mV) y el potencial de umbral es de alrededor de -55 mV. Entradas sinápticas a una neurona hacen que la membrana para despolarizar o hiperpolarizar; es decir, que hacen que el potencial de membrana para subir o bajar. Los potenciales de acción se activan cuando se acumula suficiente despolarización para llevar el potencial de membrana hasta umbral. Cuando se dispara un potencial de acción, el potencial de membrana bruscamente dispara hacia arriba, alcanzando a menudo tan alta como 100 mV, entonces igualmente bruscamente dispara de nuevo hacia abajo, terminando a menudo por debajo del nivel de reposo, donde permanece durante algún periodo de tiempo. La forma del potencial de acción es estereotipada; es decir, la subida y bajada general tienen aproximadamente la misma amplitud y curso temporal para todos los potenciales de acción en una célula dada. (Las excepciones se describen más adelante en el artículo.) En la mayoría de las neuronas, todo el proceso se lleva a cabo en menos de una milésima de segundo. Muchos tipos de neuronas emiten los potenciales de acción constantemente a tasas de hasta 10-100 por segundo; algunos tipos, sin embargo, son mucho más silenciosos, y pueden ir por minutos o más sin emitir potenciales de acción.
Base biofísica
Los potenciales de acción son el resultado de la presencia en la membrana de una célula de tipos especiales de canales iónicos dependientes de voltaje. Un canal iónico dependiente de voltaje es un grupo de proteínas incrustadas en la membrana que tiene tres propiedades fundamentales:
- Es capaz de asumir más de una conformación.
- Al menos una de las conformaciones crea un canal a través de la membrana que es permeable a los tipos específicos de iones.
- La transición entre conformaciones está influenciado por el potencial de membrana.
Así, un canal iónico dependiente de voltaje tiende a ser abierta para algunos valores de potencial de membrana, y cerrado por los demás. En la mayoría de casos, sin embargo, la relación entre el potencial de membrana y de estado de canal es probabilística e implica un retardo de tiempo. Los canales iónicos cambiar entre conformaciones en momentos impredecibles: El potencial de membrana determina la tasa de transiciones y la probabilidad por unidad de tiempo de cada tipo de transición.
Canales iónicos dependientes de voltaje son capaces de producir potenciales de acción, ya que pueden dar lugar a bucles de retroalimentación positiva: El potencial de membrana controla el estado de los canales de iones, pero el estado de los canales de iones controla el potencial de membrana. Por lo tanto, en algunas situaciones, un aumento en el potencial de membrana puede causar canales iónicos se abran, causando así un aumento adicional en el potencial de membrana. Un potencial de acción se produce cuando este ciclo de retroalimentación positiva procede de forma explosiva. El tiempo y la trayectoria de amplitud del potencial de acción están determinados por las propiedades biofísicas de los canales iónicos dependientes de voltaje que la producen. Existen varios tipos de canales que son capaces de producir la respuesta positiva necesaria para generar un potencial de acción existen. Los canales de sodio dependientes de voltaje son responsables de los potenciales de acción rápida que intervienen en la conducción nerviosa. Más lentas potenciales de acción en las células musculares y algunos tipos de neuronas son generadas por los canales de calcio dependientes de voltaje. Cada uno de estos tipos se presenta en múltiples variantes, con una sensibilidad diferente voltaje y diferentes dinámicas temporales.
El tipo más intensamente estudiado de los canales iónicos dependientes de voltaje comprende los canales de sodio que intervienen en la conducción nerviosa rápida. Estos se conocen como los canales de sodio de Hodgkin-Huxley porque se caracterizaron por primera vez por Alan Hodgkin y Andrew Huxley en su estudio de la biofísica de la acción potencial ganador del Premio Nobel, pero más convenientemente se puede denominar como canales de Na V. (La "V" es sinónimo de "tensión"). Un canal de Na V tiene tres posibles estados, conocidos como desactivado, activado y desactivado. El canal es permeable sólo a los iones sodio cuando se encuentra en el estado activado. Cuando el potencial de membrana es baja, el canal pasa la mayor parte de su tiempo en el estado desactivado (cerrado). Si el potencial de membrana se eleva por encima de un cierto nivel, el canal muestra el aumento de probabilidad de transición al estado (abierto) activado. Cuanto mayor sea el potencial de membrana mayor será la probabilidad de activación. Una vez que se ha activado un canal, con el tiempo la transición al estado inactivado (cerrado). Se tiende entonces a permanecer inactiva durante algún tiempo, pero, si el potencial de membrana se vuelve baja de nuevo, el canal eventualmente transición de vuelta al estado desactivado. Durante un potencial de acción, la mayoría de los canales de este tipo pasan por un ciclo desactivado → Activado → → inactivados desactivado. Esto es sólo el comportamiento promedio de la población, sin embargo - un canal individual puede hacer que, en principio, cualquier transición en cualquier momento. Sin embargo, la probabilidad de transición de un canal desde el estado inactivado directamente al estado activado es muy bajo: Un canal en el estado inactivado es refractario hasta que se ha hecho la transición de vuelta al estado desactivado.
El resultado de todo esto es que la cinética de los canales de Na V se rigen por una matriz de transición cuyas tasas son dependiente de la tensión de una manera complicada. Dado que estos mismos canales juegan un papel importante en la determinación de la tensión, la dinámica global del sistema puede ser bastante difícil de resolver. Hodgkin y Huxley se acercaron al problema mediante el desarrollo de un conjunto de ecuaciones diferenciales para los parámetros que rigen los estados de canal de iones, conocido como el Ecuaciones de Hodgkin-Huxley. Estas ecuaciones han sido ampliamente modificada por la investigación posterior, pero el punto de partida para la mayoría de los estudios teóricos de la biofísica potenciales de acción.
A medida que aumenta el potencial de membrana, canales iónicos de sodio abiertos, permitiendo la entrada de sodio iones en la célula. Esto es seguido por la apertura de canales iónicos de potasio que permiten la salida de potasio iones a partir de la célula. El flujo hacia el interior de iones de sodio aumenta la concentración de carga positiva cationes en la célula y causa la despolarización, donde el potencial de la celda es mayor que la celda de potencial de reposo. Los canales de sodio se cierran en el pico del potencial de acción, mientras que el potasio sigue dejando a la célula. El flujo de salida de iones potasio disminuye el potencial de membrana o hiperpolariza la célula. Para los pequeños aumentos de tensión de reposo, la corriente de potasio supera la corriente de sodio y la tensión vuelve a su valor normal en reposo, por lo general -70 mV. Sin embargo, si el voltaje aumenta más allá de un umbral crítico, típicamente 15 mV más alto que el valor de reposo, la corriente de sodio domina. Esto da lugar a una condición fuera de control mediante el cual el comentarios positivos de la corriente de sodio activa aún más los canales de sodio. Por lo tanto, la célula "fuegos", que producen un potencial de acción.
Las corrientes producidas por la apertura de los canales dependientes de voltaje en el curso de un potencial de acción son típicamente significativamente mayor que la corriente inicial estimulante. Por lo tanto, la amplitud, la duración, y la forma del potencial de acción se determinan en gran medida por las propiedades de la membrana excitable y no la amplitud o la duración del estímulo. Este propiedad de todo o nada del potencial de acción que lo diferencia de los potenciales graduados tales como los potenciales receptores, potenciales electrotonic, y potenciales sinápticos, que escala con la magnitud del estímulo. Una variedad de tipos de acción posibles existe en muchos tipos de células y compartimentos celulares que determine los tipos de canales dependientes de voltaje, canales de fugas, distribuciones de canal, concentraciones iónicas, capacitancia de la membrana, la temperatura, y otros factores.
Los principales iones que participan en un potencial de acción son cationes de sodio y potasio; iones de sodio entran en la célula, y los iones de potasio salgan, restablecer el equilibrio. Relativamente pocos iones deben atravesar la membrana para el voltaje de la membrana para cambiar drásticamente. Los iones intercambiados durante un potencial de acción, por lo tanto, hacer un cambio insignificante en las concentraciones iónicas interiores y exteriores. Los pocos iones que hacen cruz son bombeados de nuevo por la acción continua de la bomba de sodio-potasio, que, con otra transportadores de iones, mantiene la relación normal de las concentraciones de iones a través de la membrana. calcio y cationes cloruro de aniones están involucrados en algunos tipos de potenciales de acción, como la potencial de acción cardiaco y el potencial de acción en el de una sola célula alga Acetabularia, respectivamente.
Aunque los potenciales de acción se generan localmente en parches de membrana excitable, las corrientes resultantes pueden desencadenar potenciales de acción en tramos de membrana vecina, precipitando una propagación en el dominó. En contraste con difusión pasiva de potenciales eléctricos ( electrotonic potencial), los potenciales de acción se genera tramos de nuevo a lo largo excitables de la membrana y se propagan sin caries. Secciones mielinizadas de axones no son excitables y no producen potenciales de acción y la señal se propaga de forma pasiva como electrotonic potencial. Regularmente espaciados parches sin mielina, llamada nodos de Ranvier, generan potenciales de acción para aumentar la señal. Conocido como conducción saltatoria, este tipo de propagación de la señal proporciona un equilibrio favorable de la velocidad de la señal y diámetro del axón. La despolarización de terminales de los axones, en general, provoca la liberación de de neurotransmisores en la hendidura sináptica . Además, backpropagating potenciales de acción se han registrado en las dendritas de neuronas piramidales, que son omnipresentes en el neocortex. Estos se cree que tienen un papel en la pico-tiempo-dependiente plasticidad.
Neurotransmisión
Anatomía de una neurona
| Neurona |
|---|
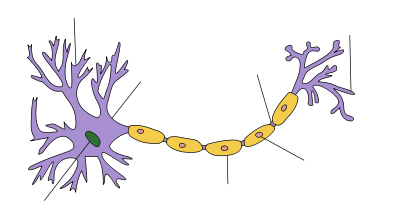 Dendrite Soma Axon Núcleo Nodo de Ranvier Terminal del axón Células de Schwann Vaina de mielina |
Varios tipos de células de soporte un potencial de acción, tales como células vegetales, células musculares, y las células especializadas del corazón (en el que se produce la potencial de acción cardiaco). Sin embargo, la célula excitable principal es la neurona, que también tiene el mecanismo más simple para el potencial de acción.
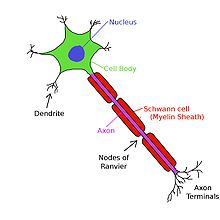
Las neuronas son células eléctricamente excitables compuestas, en general, de uno o más dendritas, un solo soma, un solo axón y uno o más terminales de los axones. La dendrita es uno de los dos tipos de sinapsis , siendo el otro el boutons terminal del axón. Las dendritas forman salientes en respuesta a los boutons terminal del axón. Estas protuberancias, o espinas, están diseñados para capturar los neurotransmisores liberados por la neurona presináptica. Tienen una alta concentración de los canales de ligando activado. Es, por lo tanto, aquí, donde las sinapsis de dos neuronas se comunican entre sí. Estas espinas tienen un cuello delgado que conecta una protuberancia bulbosa de la dendrita principal. Esto garantiza que los cambios que se producen dentro de la columna vertebral son menos propensos a afectar a las espinas vecinos. La espina dendrítica puede, por lo tanto, con raras excepciones (ver LTP), actúan como una unidad independiente. Las dendritas se conectan a la soma. Las casas de los soma núcleo, que actúa como el regulador de la neurona. A diferencia de las espinas, la superficie de la soma está poblada por tensión activado los canales iónicos. Estos canales ayudan a transmitir las señales generadas por las dendritas. Emergiendo desde el soma es la axón. Esta región se caracteriza por tener una muy alta concentración de los canales de sodio activados por voltaje. En general, se considera que es la zona de iniciación de la espiga de potenciales de acción. Múltiples señales generadas en las espinas, y se transmiten por el soma convergen aquí. Inmediatamente después del axón es la axón. Esta es una protuberancia tubular delgada viajar lejos del soma. El axón está aislado por una vaina de mielina. La mielina se compone de cualquiera Células de Schwann (en el sistema nervioso periférico) o oligodendrocitos (en el sistema nervioso central), los tipos de las células gliales. Aunque las células gliales no están involucrados con la transmisión de señales eléctricas, se comunican y proporcionan un importante apoyo bioquímica a las neuronas. Para ser más específicos, la mielina envuelve varias veces alrededor del segmento axonal, la formación de una capa de grasa gruesa que impide que los iones de entrar o escapar del axón. Este aislamiento previene la caries señal significativa, así como garantizar la velocidad de la señal más rápido. Este aislamiento, sin embargo, tiene la restricción de que no hay canales pueden estar presentes en la superficie del axón. Hay, por lo tanto, los parches regularmente espaciados de membrana, que no tienen aislamiento. Estos nodos de Ranvier pueden ser considerados como "mini lomas axón", ya que su propósito es aumentar la señal con el fin de prevenir la caries señal significativa. En el extremo más lejano, el axón pierde su aislamiento y comienza a ramificarse en varias terminales de los axones. Estos terminales de los axones luego terminan en la forma de la segunda clase de sinapsis, botones terminales del axón. Estos botones tienen canales de calcio activados por voltaje, que entran en juego cuando la señalización otras neuronas.
Iniciación
Antes de considerar la propagación de potenciales de acción a lo largo de axones y su terminación en las perillas sinápticas, es útil tener en cuenta los métodos por los que los potenciales de acción puede iniciarse en el axón. El requisito básico es que el voltaje de la membrana en el montículo se eleva por encima del umbral para el disparo. Hay varias maneras en que puede ocurrir esta despolarización.

Dinámica
Los potenciales de acción son los más iniciados por los potenciales postsinápticos excitatorios desde una neurona presináptica. Típicamente, moléculas de neurotransmisores son liberados por la presináptica neurona. Estos neurotransmisores continuación, se unen a receptores en la célula postsináptica. Esta unión se abre varios tipos de canales iónicos. Esta abertura tiene el efecto adicional de cambio de la permeabilidad local de la la membrana celular y, por lo tanto, el potencial de membrana. Si la unión aumenta la tensión (despolariza la membrana), la sinapsis excitatoria es. Si, sin embargo, la unión disminuye la tensión (hiperpolariza la membrana), es inhibitorio. Si se aumenta o disminuye la tensión, el cambio se propaga pasivamente a regiones cercanas de la membrana (como se describe por el ecuación de cable y sus refinamientos). Típicamente, el estímulo de tensión decae exponencialmente con la distancia desde la sinapsis y con el tiempo de la unión del neurotransmisor. Algunos fracción de una tensión excitadora puede llegar a la axón loma y pueden (en casos raros) despolarizar la membrana suficiente para provocar un nuevo potencial de acción. Más típicamente, los potenciales excitatorios de varias sinapsis debe trabajar juntos en casi al mismo tiempo para provocar un nuevo potencial de acción. Sus esfuerzos conjuntos pueden ser frustrados, sin embargo, por el contrarresto los potenciales postsinápticos inhibitorios.
Neurotransmisión también puede ocurrir a través de sinapsis eléctricas. Debido a la conexión directa entre las células excitables en la forma de gap junctions, un potencial de acción se pueden transmitir directamente de una célula a la siguiente en cualquier dirección. El libre flujo de iones entre las células permite la transmisión no química mediada rápida. Canales rectificadores de asegurar que los potenciales de acción se mueven en una sola dirección a través de una sinapsis eléctrica. Sinapsis eléctricas se encuentran en todos los sistemas nervioso, incluyendo el cerebro humano, a pesar de que son una clara minoría.
Principio "Todo o nada"
La amplitud de un potencial de acción es independiente de la cantidad de corriente que lo produjo. En otras palabras, las corrientes más grandes no crean potenciales de acción más grandes. Por lo tanto, los potenciales de acción se dice que son señales de todo o nada, ya sea que se produzcan en todo o que no se producen en absoluto. La frecuencia de potenciales de acción se correlaciona con la intensidad de un estímulo. Esto está en contraste con los potenciales receptores, cuyas amplitudes dependen de la intensidad de un estímulo.
Las neuronas sensoriales
En las neuronas sensoriales, una señal externa, como la presión, la temperatura, la luz o el sonido se acopla con la apertura y cierre de canales iónicos, que a su vez alteran las permeabilidades iónicos de la membrana y su tensión. Estos cambios de tensión pueden volver a ser excitatorio (despolarizante) o inhibidor (hiperpolarización) y, en algunos neuronas sensoriales, sus efectos combinados pueden despolarizar el axón suficiente para provocar potenciales de acción. Ejemplos en los seres humanos incluyen la neurona receptor olfativo y Corpúsculo de Meissner, que son fundamentales para el sentido del olor y tocar, respectivamente. Sin embargo, no todas las neuronas sensoriales convertir sus señales externas en los potenciales de acción; algunos ni siquiera tienen un axón! En cambio, pueden convertir la señal en el lanzamiento de un neurotransmisor, o en continuo potenciales calificados, cualquiera de los cuales pueden estimular posterior neurona (s) en el despido de un potencial de acción. Por ejemplo, en el ser humano oído, células ciliadas convierten el sonido entrante en la apertura y cierre de mecánicamente canales iónicos, que puede causar moléculas de neurotransmisores que se liberan. En forma similar, en el ser humano retina , la inicial células fotorreceptoras y la siguiente capa de células (que comprende células bipolares y células horizontales) no producen potenciales de acción; solo algunos células amacrinas y la tercera capa, la las células ganglionares, producen potenciales de acción, que luego viajar hasta el nervio óptico.
Potenciales marcapasos

En las neuronas sensoriales, los potenciales de acción son el resultado de un estímulo externo. Sin embargo, algunas células excitables no requieren este tipo de estímulo a fuego: Espontáneamente despolarizan sus potenciales axón y la acción del fuego a un ritmo regular, como un reloj interno. Las trazas de tensión de tales células se conocen como potenciales marcapasos. La células marcapasos cardíacos del nodo sinusal en el corazón son un buen ejemplo. Aunque dichos potenciales marcapasos tienen una ritmo natural, que se puede ajustar por estímulos externos; por ejemplo, ritmo cardíaco puede ser alterado por los productos farmacéuticos, así como las señales de la simpático y nervios parasimpáticos. Los estímulos externos no causan disparo repetitivo de la célula, sino que simplemente alteran su temporización. En algunos casos, la regulación de la frecuencia puede ser más complejo, dando lugar a patrones de potenciales de acción, tales como reventar.
Fases
El curso del potencial de acción se puede dividir en cinco partes: la fase de ascenso, la fase pico, la fase de caída, la fase de aterrizaje corto, y el período refractario. Durante la fase de aumento de los potenciales de membrana se despolariza (se vuelve más positivo). El punto en que se detiene la despolarización se llama la fase pico. En esta etapa, el potencial de membrana alcanza un máximo. Con posterioridad a esto, hay una fase de caída. Durante esta etapa los hiperpolariza potenciales de membrana (se hace más negativa). La fase inferior es el punto en el que el potencial de membrana se convierte temporalmente cargada más negativamente que cuando está en reposo. Finalmente, el tiempo durante el cual un potencial de acción posterior es imposible o difícil de fuego se llama período refractario, que puede solaparse con las otras fases.
El curso del potencial de acción se determina por dos efectos acoplados. En primer lugar, los canales iónicos sensibles al voltaje se abren y cierran en respuesta a cambios en el voltaje de la membrana V m. Esto cambia la permeabilidad de la membrana a los iones. En segundo lugar, de acuerdo con la Ecuación de Goldman, este cambio en los cambios de permeabilidad en el potencial de equilibrio E m, y, por tanto, el voltaje de la membrana V m. Por lo tanto, el potencial de membrana afecta a la permeabilidad, que luego afecta aún más el potencial de membrana. Esto establece la posibilidad de que regeneración positiva, que es una parte clave de la fase ascendente del potencial de acción. Un factor que complica es que un único canal iónico puede tener varias "puertas" internas que responden a cambios en V m de maneras opuestas, o a diferentes velocidades. Por ejemplo, aunque el aumento V m abre la mayoría de las puertas en el canal de sodio sensibles al voltaje, también se cierra "compuerta de inactivación" del canal, aunque más lentamente. Por lo tanto, cuando V m se eleva de repente, los canales de sodio se abren inicialmente, pero a continuación, cierre debido a la inactivación lenta.
Los voltajes y las corrientes del potencial de acción en todas sus fases fueron modelados con precisión por Alan Lloyd Hodgkin y Andrew Huxley en 1952, para el que fueron galardonados con el Premio Nobel de Fisiología o Medicina en 1963. Sin embargo, su modelo considera sólo dos tipos de canales iónicos sensibles al voltaje, y hace varias suposiciones acerca de ellos, por ejemplo, que sus puertas internas se abren y cierran de forma independiente el uno del otro. En realidad, hay muchos tipos de canales iónicos, y no siempre se abren y cierran de forma independiente.
La estimulación y el aumento de fase
Un potencial de acción típico comienza en el axón con una despolarización suficientemente fuerte, por ejemplo, un estímulo que aumenta V m. Esta despolarización es a menudo causada por la inyección de sodio adicionales cationes en la célula; estos cationes pueden provenir de una amplia variedad de fuentes, tales como sinapsis químicas , neuronas sensoriales o potenciales marcapasos.
Para una neurona en reposo, existe una alta concentración de iones de sodio y cloro en la líquido extracelular en comparación con el líquido intracelular, mientras que hay una alta concentración de iones de potasio en el líquido intracelular en comparación con el líquido extracelular. Este gradiente de concentración junto con canales de fugas de potasio presentes en la membrana de la neurona provoca una flujo de salida de iones de potasio que hacen el potencial de reposo cerca de E K ≈ -75 mV. La despolarización se abre tanto el sodio y los canales de potasio en la membrana, permitiendo que los iones fluyan dentro y fuera del axón, respectivamente. Si la despolarización es pequeño (por ejemplo, el aumento de V m de -70 mV a -60 mV), la corriente de salida de potasio supera la corriente de sodio hacia el interior y la membrana repolariza de nuevo a su potencial normal de reposo alrededor de -70 mV. Sin embargo, si la despolarización es lo suficientemente grande, las sodio hacia el interior aumenta la corriente superior a la corriente de salida de potasio y una condición fuera de control ( retroalimentación) resultados positivos: el más corriente de entrada hay, más V m aumenta, lo que a su vez aumenta más la corriente de entrada. A lo suficientemente fuerte despolarización (aumento en V m) provoca que los canales de sodio sensibles al voltaje para abrir; el aumento de la permeabilidad al sodio conduce V m más cerca de la tensión de equilibrio de sodio Na E ≈ 55 mV. La tensión creciente a su vez provoca aún más los canales de sodio para abrir, que empuja V m aún más hacia E Na. Esta retroalimentación positiva continúa hasta que los canales de sodio están completamente abiertas y V m es cerca de E Na. El fuerte aumento de V m y la permeabilidad de sodio corresponden a la fase ascendente del potencial de acción.
La tensión de umbral crítico para esta condición fuera de control es generalmente alrededor de -45 mV, pero depende de la actividad reciente del axón. Una membrana que se acaba de disparado un potencial de acción no puede disparar otro inmediatamente, ya que los canales iónicos no han vuelto a su estado habitual. El período durante el cual no nuevo potencial de acción puede ser despedido se denomina período refractario absoluto. En tiempos más largos, después de algunos, pero no todos los canales de iones se han recuperado, el axón puede ser estimulado para producir otro potencial de acción, pero sólo con una despolarización mucho más fuerte, por ejemplo, -30 mV.El período durante el cual los potenciales de acción son inusualmente difíciles de evocar es llamado elperíodo refractario relativo.
Pico y la caída de fase
La retroalimentación positiva de la fase ascendente se ralentiza y se detiene como los canales de iones de sodio se vuelven de máxima abertura. En el pico del potencial de acción, la permeabilidad de sodio y se maximiza el voltaje de la membrana V m es casi igual a la tensión de equilibrio de sodio E N / A . Sin embargo, la misma tensión elevada que abre los canales de sodio inicialmente también les cierra lentamente, cerrando sus poros; se convierten en los canales de sodio inactivados . Esto reduce la permeabilidad de la membrana al relativa de sodio a potasio, conduciendo el voltaje de la membrana hacia el valor de reposo. Al mismo tiempo, el voltaje elevado abre los canales de potasio sensibles al voltaje; el aumento de la permeabilidad de las unidades de potasio de la membrana V m hacia E K . En conjunto, estos cambios en sodio y potasio causa permeabilidad V m a caer rápidamente, repolarización de la membrana y la producción de la "caída de fase" del potencial de acción.
Afterhyperpolarization
La tensión elevada abrió muchos más canales de potasio que de costumbre, y algunos de ellos no se cierran de inmediato cuando la membrana vuelve a su tensión normal de reposo. Adicionalmente, más canales de potasio se abren en respuesta a la afluencia de iones de calcio durante el potencial de acción. La permeabilidad de potasio de la membrana es transitoriamente inusualmente alta, la conducción del voltaje de la membrana V m aún más cerca de la tensión de equilibrio de potasio E K . Por lo tanto, hay una undershoot o hiperpolarización, denominado un afterhyperpolarization en lenguaje técnico, que persiste hasta la membrana de permeabilidad de potasio vuelve a su valor habitual.
Periodo refractario
Cada potencial de acción es seguido por un período refractario, que puede ser dividido en un periodo refractario absoluto , durante el cual es imposible para evocar otro potencial de acción, y luego un período refractario relativo , durante el cual se requiere un estímulo más fuerte de lo habitual. Estos dos períodos refractarios son causados por cambios en el estado de las moléculas de canales de sodio y de potasio. Cuando se cierra después de un potencial de acción, los canales de sodio entran en un estado "inactivada", en el que no se pueden hacer para abrir independientemente del potencial de membrana-esto da lugar a el periodo refractario absoluto. Incluso después de un número suficiente de canales de sodio han hecho la transición de nuevo a su estado de reposo, sucede con frecuencia que una fracción de los canales de potasio permanece abierta, por lo que es difícil para el potencial de membrana para despolarizar, y dando lugar así a el período refractario relativo. Debido a que la densidad y subtipos de canales de potasio pueden diferir mucho entre los diferentes tipos de neuronas, la duración del período refractario relativo es muy variable.
Una vez que un potencial de acción se ha producido en un parche de membrana, el parche de membrana necesita tiempo para recuperarse antes de que pueda disparar de nuevo. A nivel molecular, este periodo refractario absoluto corresponde al tiempo requerido para los canales de sodio activados por voltaje para recuperarse de inactivación, es decir, para volver a su estado cerrado. Hay muchos tipos de canales de potasio activados por voltaje en las neuronas, algunos de ellos inactivar (corrientes de tipo A) rápidas y algunos de ellos inactivan lentamente o no inactivar en absoluto; esta variabilidad garantiza que habrá siempre una fuente disponible de la corriente de repolarización, incluso si algunos de los canales de potasio son inactivados por la despolarización anterior. Por otro lado, todos los canales de sodio neuronales activados por voltaje inactivan dentro de varios milisegundos durante fuerte despolarización, haciendo así imposible siguiente despolarización hasta que una fracción sustancial de los canales de sodio han vuelto a su estado cerrado. A pesar de que limita la frecuencia de disparo, el período refractario absoluto asegura que los potenciales de acción se mueve en una sola dirección a lo largo de un axón. Las corrientes que fluyen en debido a un potencial de acción extendido en ambas direcciones a lo largo del axón. Sin embargo, sólo la parte no disparada del axón puede responder con un potencial de acción; la parte que acaba despedido no responde hasta que el potencial de acción es con seguridad fuera de alcance y no puede reestimular esa parte. En el habitual de conducción ortodrómica, el potencial de acción se propaga desde la loma axón hacia las perillas sinápticas (el termini axonal); propagación en la dirección opuesta conocido como antidrómica conducción-es muy raro. Sin embargo, si un axón laboratorio se estimula en su parte media, las dos mitades del axón son "fresco", es decir, sin cocer; a continuación, se generarán dos potenciales de acción, uno que viaja hacia el cono axónico y la otra viaja hacia las perillas sinápticas.
Mielina y conducción relativa a la danza
A fin de permitir la transducción de rápido y eficiente de las señales eléctricas en el sistema nervioso, ciertos axones neuronales están cubiertas con vainas de mielina. La mielina es una membrana multilamelares que envuelve el axón en segmentos separados por intervalos conocidos como nodos de Ranvier. Es producida por células especializadas: las células de Schwann exclusivamente en el sistema nervioso periférico, y oligodendrocitos exclusivamente en la sistema nervioso central. vaina de mielina reduce capacitancia de la membrana y aumenta la resistencia de la membrana en los intervalos inter-nodo, permitiendo así un movimiento rápido, saltatoria de potenciales de acción de nodo a nodo. La mielinización se encuentra principalmente en los vertebrados , pero un sistema análogo se ha descubierto en un par de invertebrados, tales como algunas especies de camarón. No todas las neuronas de vertebrados son mielinizadas; por ejemplo, los axones de las neuronas que constituyen el sistema nervioso autónomo no son, en general, mielinizadas.
La mielina impide que los iones entren o salgan del axón largo de los segmentos mielinizados. Como regla general, la mielinización aumenta la velocidad de conducción de los potenciales de acción y los hace más eficientes energéticamente. Ya sea saltatoria o no, la velocidad media de conducción de un potencial de acción varía de 1 m / s a más de 100 m / s, y, en general, aumenta con el diámetro axonal.
Los potenciales de acción no pueden propagarse a través de la membrana en segmentos mielinizados del axón. Sin embargo, la corriente es transportada por el citoplasma, que es suficiente para despolarizar la primera o segunda subsiguiente nodo de Ranvier. En lugar de ello, la corriente iónica a partir de un potencial de acción en un nodo de Ranvier provoca otro potencial de acción en el siguiente nodo; esta aparente "saltos" del potencial de acción de nodo a nodo que se conoce como la conducción relativa a la danza. Aunque el mecanismo de conducción saltatoria se sugirió en 1925 por Ralph Lillie, la primera evidencia experimental de conducción saltatoria vino de Ichiji Tasaki y Taiji Takeuchi y de Andrew Huxley y Robert Stämpfli. Por el contrario, en los axones no mielinizados, el potencial de acción provoca otro en la membrana inmediatamente adyacente, y se mueve continuamente por el axón como una ola.
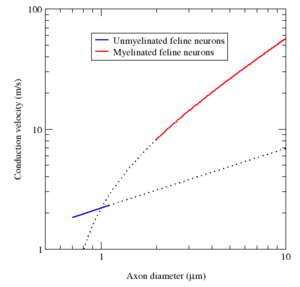
La mielina tiene dos ventajas importantes: la velocidad de conducción rápido y la eficiencia energética. Para los axones de mayor tamaño que un diámetro mínimo (aproximadamente 1 micrómetro), la mielinización aumenta la velocidad de conducción de un potencial de acción, típicamente diez veces. A la inversa, para una velocidad de conducción dado, fibras mielinizadas son más pequeños que sus homólogos no mielinizadas. Por ejemplo, los potenciales de acción mover aproximadamente a la misma velocidad (25 m / s) en un axón rana mielinizadas y amielínicas un axón gigante del calamar, pero el axón rana tiene una más o menos 30 veces más pequeño diámetro y 1.000 veces el área de la sección transversal más pequeña . Además, dado que las corrientes iónicas se limitan a los nodos de Ranvier, muchos menos iones "fugas" a través de la membrana, el ahorro de energía metabólica. Este ahorro es una significativa ventaja selectiva , ya que el sistema nervioso humano utiliza aproximadamente 20% de la energía metabólica del cuerpo.
La longitud de los segmentos mielinizados axones 'es importante para el éxito de la conducción relativa a la danza. Ellos deben ser tan largo como sea posible para maximizar la velocidad de conducción, pero no tan largo que la señal que llega es demasiado débil para provocar un potencial de acción en el siguiente nodo de Ranvier. En la naturaleza, los segmentos mielinizados son generalmente lo suficiente para que la señal propagada pasivamente a viajar por al menos dos nodos al tiempo que conserva suficiente amplitud para disparar un potencial de acción en el segundo o tercer nodo. Por lo tanto, el factor de seguridad de la conducción saltatoria es alta, lo que permite la transmisión a los nodos de derivación en caso de lesión. Sin embargo, los potenciales de acción pueden terminar prematuramente en ciertos lugares donde el factor de seguridad es bajo, incluso en neuronas amielínicas; Un ejemplo común es el punto de un axón, donde se divide en dos axones rama.
Algunas enfermedades degradan la mielina y perjudican la conducción relativa a la danza, la reducción de la velocidad de conducción de los potenciales de acción. El bien conocido de estos es más esclerosis múltiple , en el que la descomposición de la mielina afecta movimiento coordinado.
Teoría de cable
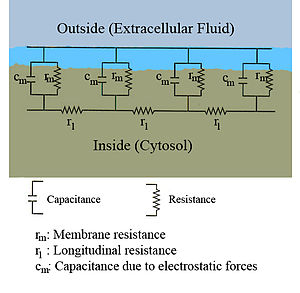
El flujo de las corrientes dentro de un axón se puede describir cuantitativamente mediante la teoría de cable y sus elaboraciones, como el modelo compartimental. Teoría cable fue desarrollado en 1855 por Lord Kelvin para modelar el cable telegráfico transatlántico y ha demostrado ser relevante para las neuronas de Hodgkin y Rushton en 1946. En teoría simple cable, la neurona es tratado como un cable de transmisión perfectamente cilíndrica eléctricamente pasiva, que puede ser descrito por una ecuación diferencial parcial

donde V (x, t) es el voltaje a través de la membrana en un momento t y una posición x a lo largo de la longitud de la neurona, y donde λ y τ son las escalas de longitud y de tiempo característicos en que dichas tensiones de decaimiento en respuesta a un estímulo . Haciendo referencia al diagrama de circuito anterior, estas escalas se pueden determinar a partir de las resistencias y capacitancias por unidad de longitud

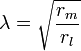
Estos hora y duración escalas se pueden utilizar para comprender la dependencia de la velocidad de conducción en el diámetro de la neurona en fibras no mielinizadas. Por ejemplo, la escala de tiempo τ-aumenta tanto con la resistencia de la membrana r m y la capacitancia c m . Como la capacitancia aumenta, más carga debe ser transferido para producir un voltaje transmembrana dado (por la ecuación Q = CV ); como la resistencia aumenta, menos carga se transfiere por unidad de tiempo, haciendo que el equilibrio más lento. En forma similar, si la resistencia interna por unidad de longitud r yo es menor en un axón que en otro (por ejemplo, porque el radio de la primera es más grande), la longitud espacial λ caries se vuelve más largo y la velocidad de conducción de un potencial de acción debe aumentar . Si la resistencia transmembrana r m se incrementa, que disminuye la corriente media "fuga" a través de la membrana, del mismo modo causando λ para convertirse en más tiempo, aumentando la velocidad de conducción.
Terminación
Sinapsis químicas
En general, los potenciales de acción que llegan a los mandos sinápticas causan un neurotransmisor que se libera en la hendidura sináptica. Los neurotransmisores son moléculas pequeñas que pueden abrir los canales de iones en la célula postsináptica; la mayoría de los axones tienen el mismo neurotransmisor en todos sus extremos. La llegada del potencial de acción abre los canales de calcio sensibles al voltaje en la membrana presináptica; la afluencia de calcio causa vesículas llenas de neurotransmisor a migrar a la superficie de la célula y liberar su contenido en la hendidura sináptica . Este complejo proceso se inhibe por las neurotoxinas tetanospasmina y toxina botulínica, que son responsables por el tétanos y el botulismo, respectivamente.

Sinapsis eléctricas
Algunas sinapsis dispensan con el "intermediario" del neurotransmisor, y conectan las células presinápticas y postsinápticas juntos. Cuando un potencial de acción alcanza una sinapsis tales, las corrientes iónicas que fluyen en la célula presináptica pueden cruzar la barrera de las dos membranas celulares y entrar en la célula postsináptica a través de poros conocidos como connexons. Por lo tanto, las corrientes iónicas del potencial de acción presináptica pueden estimular directamente la célula postsináptica. Sinapsis eléctricas permiten una transmisión más rápida, ya que no requieren la lenta difusión de los neurotransmisores a través de la hendidura sináptica. Por lo tanto, las sinapsis eléctricas se utilizan cada vez que una respuesta rápida y la coordinación de la oportunidad son cruciales, como en los reflejos de escape, la retina de los vertebrados , y el corazón .
Uniones neuromusculares
Un caso especial de una sinapsis química es la unión neuromuscular, en la que el axón de una neurona motora termina en una fibra muscular. En tales casos, el neurotransmisor liberado es la acetilcolina, que se une al receptor de acetilcolina, una proteína integral de membrana en la membrana (el sarcolema ) de la fibra muscular. Sin embargo, la acetilcolina no permanece unido; más bien, se disocia y se hidroliza por la enzima, acetilcolinesterasa, situada en la sinapsis. Esta enzima reduce rápidamente el estímulo al músculo, que permite que el grado y la sincronización de la contracción muscular para ser regulado con delicadeza. Algunos venenos inactivan acetilcolinesterasa para evitar este control, tales como los agentes nerviosos sarín y tabun y el insecticidas diazinón y malatión.
Otros tipos de células
Los potenciales de acción cardiacos

El potencial de acción cardíaco difiere del potencial de acción neuronal por tener una meseta extendida, en la que la membrana se mantiene a una alta tensión para unos pocos cientos de milisegundos antes de ser repolarized por la corriente de potasio como de costumbre. Esta meseta es debido a la acción de más lentos de calcio canales de apertura y que sujetan el voltaje de la membrana cerca de su potencial de equilibrio, incluso después de los canales de sodio han inactivado.
El potencial de acción cardíaca desempeña un papel importante en la coordinación de la contracción del corazón. Las células cardíacas del nodo sinoauricular proporcionan el potencial marcapasos que sincroniza el corazón. Los potenciales de acción de esas células se propagan hacia ya través del nodo atrioventricular (nodo AV), que es normalmente la única vía de conducción entre el aurículas y los ventrículos. Los potenciales de acción desde el nodo AV viajes a través del haz de His y de allí a las fibras de Purkinje. A la inversa, anomalías en el potencial de acción cardíaco, ya sea debido a una mutación congénita o lesión puede conducir a patologías humanas, especialmente arritmias. Varios fármacos antiarrítmicos actúan sobre el potencial de acción cardiaco, tales como quinidina, lidocaína, bloqueadores beta, y verapamil.
Los potenciales de acción muscular
El potencial de acción en una célula de músculo esquelético normal es similar a la del potencial de acción en las neuronas. Los potenciales de acción resultan de la despolarización de la membrana celular (el sarcolema), que abre los canales de sodio sensibles al voltaje; estos se convierten en inactivado y la membrana es repolarizadas a través de la corriente hacia el exterior de iones de potasio. El potencial de reposo antes de la potencial de acción es típicamente -90mV, algo más negativo que las neuronas típicas. El potencial de acción muscular dura aproximadamente 2-4 ms, el periodo refractario absoluto es de aproximadamente 1-3 ms, y la velocidad de conducción a lo largo del músculo es de aproximadamente 5 m / s. El potencial de acción libera calcio iones que liberan la tropomiosina y permiten que el músculo se contraiga. Los potenciales de acción muscular son provocados por la llegada de un potencial de acción neuronal presináptica en la unión neuromuscular, que es un objetivo común para neurotoxinas.
Los potenciales de acción Plant
Planta y células de hongos también son eléctricamente excitables. La diferencia fundamental de los potenciales de acción animal es, que la despolarización en las células vegetales no se logra una absorción de iones de sodio positivos, pero por la liberación de negativos de cloruro de iones. Junto con la siguiente liberación de los iones de potasio positivos, que es común a los potenciales de acción de plantas y animales, el potencial de acción en plantas deduce, por lo tanto, una pérdida osmótica de sal (KCl), mientras que el potencial de acción animal es osmóticamente neutral, cuando cantidades iguales de entrar en sodio y potasio dejando anulan entre sí osmóticamente. La interacción de las relaciones eléctricas y osmóticos en células vegetales indica una función osmótica de la excitabilidad eléctrica de los ancestros comunes, unicelulares de plantas y animales en condiciones de salinidad cambiante, mientras que la presente función de transmisión de la señal rápida es visto como un logro menor de metazoos células en un entorno osmótica más estable. Hay que suponer que la función de señalización familiar de potenciales de acción en algunas plantas vasculares (por ejemplo, Mimosa pudica ), surgió independientemente de que en las células excitables metazoos.
Distribución taxonómica y ventajas evolutivas
Los potenciales de acción se encuentran en todo los organismos multicelulares, incluyendo plantas , invertebrados tales como insectos y vertebrados como reptiles y mamíferos . Esponjas parece ser el principal phylum de multicelulares eucariotas que no transmiten los potenciales de acción, aunque algunos estudios han sugerido que estos organismos tienen una forma de la señalización eléctrica, también. El potencial de reposo, así como el tamaño y la duración del potencial de acción, no han variado mucho con la evolución, aunque la velocidad de conducción varía dramáticamente con diámetro axonal y la mielinización.
| Animal | Tipo de la célula | Potencial de reposo (mV) | Aumento AP (mV) | AP duración (ms) | Velocidad de conducción (m / s) |
|---|---|---|---|---|---|
| Calamar (Loligo) | Axón gigante | -60 | 120 | 0.75 | 35 |
| Lombriz de tierra (Lumbricus) | La mediana de fibra gigante | -70 | 100 | 1.0 | 30 |
| Cucaracha (Periplaneta) | Fibra gigante | -70 | 80-104 | 0.4 | 10 |
| Frog (Rana) | Axón del nervio ciático | -60--80 | 110-130 | 1.0 | 7-30 |
| Gato (Felis) | Motoneurona espinal | -55--80 | 80-110 | 1-1,5 | 30-120 |
Dada su conservación a lo largo de la evolución, el potencial de acción parece conferir ventajas evolutivas. Una de las funciones de los potenciales de acción es rápida, la señalización de largo alcance dentro del organismo; la velocidad de conducción puede exceder de 110 m / s, que es un tercio de la velocidad del sonido. de comparación, una molécula de hormona realizado en el torrente sanguíneo se mueve a aproximadamente 8 m / s en las grandes arterias. Parte de esta función es la estrecha coordinación de eventos mecánicos, tales como la contracción del corazón. Una segunda función es el cálculo asociado con su generación. Al ser una señal de todo o nada que no decae con la distancia de transmisión, el potencial de acción tiene ventajas similares a la electrónica digital. La integración de varias señales dendríticas en el axón y su umbral para formar un tren complejo de potenciales de acción es otra forma de cálculo, uno que ha sido explotada biológicamente para formar generadores centrales de patrones y imitado en redes neuronales artificiales.
Métodos experimentales

El estudio de los potenciales de acción ha requerido el desarrollo de nuevos métodos experimentales. El trabajo inicial, antes de 1955, se centró en tres objetivos: señales de aislamiento de las neuronas o axones individuales, de rápido desarrollo, electrónicos sensibles, y la disminución de electrodos suficientes de que la tensión dentro de una sola célula podría ser registrada.
El primer problema se resolvió mediante el estudio de los axones gigantes que se encuentran en las neuronas del género calamar Loligo . Estos axones son tan grandes de diámetro (aproximadamente 1 mm o 100 veces más grande que una neurona típica) que se pueden ver a simple vista, haciendo que sean fáciles de extraer y manipular. Sin embargo, los Loligo axones no son representativos de todas las células excitables, y numerosos otros sistemas con los potenciales de acción se han estudiado.
El segundo problema fue abordado con el desarrollo fundamental de la fijación de voltaje, lo que permitió experimentadores para estudiar las corrientes iónicas que subyacen a un potencial de acción de forma aislada, y se elimina una importante fuente de ruido electrónico, la corriente I C asociada a la capacitancia C de la membrana. Puesto que la corriente es igual a C veces la tasa de cambio de la tensión de transmembrana V m , la solución era diseñar un circuito que mantiene V m fija (tasa cero de cambio) independientemente de las corrientes que fluyen a través de la membrana. Por lo tanto, la corriente requerida para mantener V m en un valor fijo es un reflejo directo de la corriente que fluye a través de la membrana. Otros avances electrónicos incluyen el uso de jaulas de Faraday y la electrónica con alta impedancia de entrada, de modo que la propia medición no afectó que se mide la tensión.
El tercer problema, el de la obtención de electrodos suficientemente pequeñas para tensiones de registro dentro de un solo axón sin perturbar ella, se resolvió en 1949 con la invención del electrodo de micropipeta de vidrio, que fue adoptado rápidamente por otros investigadores. Los refinamientos de este método son capaces de producir puntas de los electrodos que son tan fina como 100 Å (10 nm), que también confiere una alta impedancia de entrada. Los potenciales de acción también pueden ser grabadas con electrodos metálicos pequeños colocados justo al lado de una neurona, con neurochips que contienen EOSFETs, u ópticamente con colorantes que son sensibles a Ca 2+ o de tensión.

Mientras electrodos micropipeta de vidrio miden la suma de las corrientes que pasan a través de muchos canales de iones, que estudian las propiedades eléctricas de un solo canal de iones se convirtió en posible en la década de 1970 con el desarrollo de la patch clamp por Erwin Neher y Bert Sakmann. Para esto fueron galardonados con el Premio Nobel de Fisiología o Medicina en 1991. Patch-sujeción verificado que los canales iónicos tienen estados discretos de la conductancia, como abierto, cerrado e inactivado.
Tecnologías de formación de imágenes ópticas se han desarrollado en los últimos años para medir los potenciales de acción, ya sea a través de múltiples sitios grabaciones simultáneas o con la resolución ultra-espacial. Uso tintes sensibles al voltaje, los potenciales de acción se han registrado ópticamente de un pequeño parche demembrana de los cardiomiocitos.
Neurotoxinas

Varios neurotoxinas, tanto naturales como sintéticos, están diseñados para bloquear el potencial de acción. tetrodotoxina del pez globo y saxitoxina del Gonyaulax (el género dinoflagelado responsable de " mareas rojas ") los potenciales de acción de bloques mediante la inhibición de los canales de sodio sensibles al voltaje; del mismo modo, dendrotoxina de la serpiente mamba negro inhibe el canal de potasio sensibles al voltaje. Tales inhibidores de los canales iónicos sirven un propósito importante investigación, permitiendo a los científicos a "apagar" los canales específicos a voluntad, aislando así las aportaciones de los otros canales; también pueden ser útiles en la purificación de los canales iónicos por cromatografía de afinidad o en el ensayo de su concentración. Sin embargo, estos inhibidores también hacen neurotoxinas eficaces, y se han considerado para el uso como armas químicas. neurotoxinas dirigidas a los canales iónicos de insectos han sido eficaces los insecticidas; un ejemplo es el sintético permetrina, que prolonga la activación de los canales de sodio que participan en los potenciales de acción. Los canales iónicos de insectos son lo suficientemente diferentes de sus homólogos humanos que hay pocos efectos secundarios en los seres humanos. Muchas otras neurotoxinas interfieren con la transmisión de los efectos del potencial de acción en las sinapsis , especialmente en la unión neuromuscular.
Historia
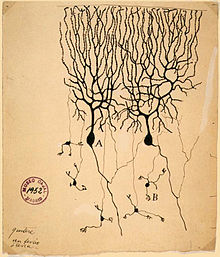
El papel de la energía eléctrica en el sistema nervioso de los animales se observó por primera vez en disecadosranasporLuigi Galvani, quien estudió desde 1791 a 1797. Los resultados de Galvani estimuladoAlessandro Volta para desarrollar lapila voltaica-la-antigua que se conocede la batería con eléctrico que estudió electricidad animal (comoanguilas eléctricas) y las respuestas fisiológicas al aplicadasde corriente continuavoltajes.
Los científicos del siglo 19a estudiaron la propagación de señales eléctricas en todo nervios (es decir, haces de neuronas) y demostraron que el tejido nervioso se compone de células , en lugar de una red interconectada de tubos (un retículo ). Carlo Matteucci siguió estudios de Galvani y demostrado que las membranas celulares tenían un voltaje a través de ellos y podrían producir corriente continua. El trabajo de Matteucci inspiró el fisiólogo alemán, Emil du Bois-Reymond, quien descubrió el potencial de acción en 1848. La velocidad de conducción de los potenciales de acción se midió por primera vez en 1850 por el amigo de du Bois-Reymond, Hermann von Helmholtz. Establecer que el tejido nervioso está formado por células discretas, el médico español Santiago Ramón y Cajal y sus estudiantes utilizan una mancha desarrollado por Camillo Golgi para revelar las formas innumerables de neuronas, que se prestan minuciosamente. Por sus descubrimientos, Golgi y Ramón y Cajal fueron galardonados con el 1906 Premio Nobel de Fisiología. Su trabajo resuelve una controversia de larga data en la neuroanatomía del siglo 19; Golgi mismo había defendido el modelo de red del sistema nervioso.
El siglo 20 fue una época dorada para electrofisiología. En 1902 y nuevamente en 1912, Julius Bernstein avanzó la hipótesis de que el potencial de acción resultado de un cambio en la permeabilidad de la membrana axonal a los iones. La hipótesis de Bernstein fue confirmada por Ken Cole y Howard Curtis, que mostró que los aumentos de conductancia de la membrana durante un potencial de acción. En 1907, Louis Lapicque sugiere que el potencial de acción se generó como fue cruzado un umbral, lo que sería más tarde muestra como un producto de los sistemas dinámicos de conductancias iónicas. En 1949, Alan Hodgkin y Bernard Katz refinado hipótesis de Bernstein al considerar que la membrana axonal podría tener diferentes permeabilidades a diferentes iones; en particular, demostraron el papel crucial de la permeabilidad de sodio durante el potencial de acción. Esta línea de investigación culminó en los cinco 1952 papeles de Hodgkin, Katz y Andrew Huxley, en la que se aplicó la técnica de fijación de voltaje para determinar la dependencia de la permeabilidad de la membrana axonal a los iones de sodio y potasio en el voltaje y el tiempo, de las que pudieron para reconstruir el potencial de acción cuantitativamente. Hodgkin y Huxley correlacionan las propiedades de su modelo matemático con discretos canales iónicos que pudieran existir en varios estados diferentes, incluyendo "abierto", "cerrado" y "inactiva". Sus hipótesis se confirmaron a mediados de los años 1970 y 1980 por Erwin Neher y Bert Sakmann, quien desarrolló la técnica de patch clamp para examinar los estados de conductancia de los canales de iones individuales. En el siglo 21, los investigadores están empezando a comprender la base estructural de estos estados de conductancia y de la selectividad de los canales por sus especies de iones a través de las atómica resolución de estructuras cristalinas, las mediciones de distancia de fluorescencia y estudios de crio-microscopía electrónica.
Julius Bernstein también fue el primero en introducir la ecuación de Nernst para el potencial a través de la membrana en reposo; este fue generalizado por David E. Goldman a la epónima ecuación de Goldman en 1943. La bomba de sodio-potasio fue identificado en 1957 y sus propiedades dilucidado gradualmente, culminando en la determinación de su estructura atómica resolución por cristalografía de rayos X. Las estructuras cristalinas de las bombas iónicas relacionados también han sido resueltos, dando una visión más amplia de cómo funcionan estas máquinas moleculares.
Los modelos cuantitativos

Los modelos matemáticos y computacionales son esenciales para la comprensión del potencial de acción, y ofrecen predicciones que pueden probarse con los datos experimentales, proporcionando una prueba rigurosa de una teoría. El más importante y precisa de estos modelos es el modelo de Hodgkin-Huxley, que describe el potencial de acción por un conjunto acoplado de cuatro ecuaciones diferenciales ordinarias (EDOs). Aunque el modelo de Hodgkin-Huxley puede ser una simplificación de una membrana nervioso realista, tal como existe en la naturaleza, su complejidad ha inspirado varios modelos incluso-más-simplificados, tales como el modelo de Morris-Lecar y el modelo Fitzhugh-Nagumo, ambos de los cuales tener sólo dos EDOs acoplados. Las propiedades de los modelos de Hodgkin-Huxley y Fitzhugh-Nagumo y sus familiares, como el modelo der Pol Bonhoeffer-van, han sido bien estudiados dentro de las matemáticas, computación y electrónica. Una investigación más moderna se ha centrado en sistemas más grandes y más integrados; uniéndose a los modelos de acción potencial con modelos de otras partes del sistema nervioso (tales como dendritas y sinapsis), las investigaciones pueden estudiar computación y simples neuronales reflejos, como los reflejos de escape y otras controladas por los generadores centrales de patrones.