

Centro de reacción fotosintética
Sabías ...
Esta selección Escuelas fue originalmente elegido por SOS para las escuelas en el mundo en desarrollo que no tienen acceso a Internet. Está disponible como una descarga intranet. Madres SOS cada aspecto después de un una familia de niños apadrinados .
Un centro de reacción fotosintética (o centro de reacción fotosintética) es un complejo de varias proteínas, pigmentos y otros co-factores ensamblados juntos para ejecutar las reacciones de conversión de energía primaria de la fotosíntesis . Excitaciones moleculares, ya sea procedente directamente de la luz solar o transferidos como energía de excitación a través de sistemas de antena captadores de luz, dan lugar a reacciones de transferencia de electrones a lo largo de una serie de cofactores unidos a proteínas. Estos cofactores son moléculas que absorben la luz (también nombrados cromóforos o pigmentos), tales como clorofila y feofitina, así como quinonas. La energía del fotón se utiliza para promover un electrón a una más alto nivel de energía molecular de un pigmento. La energía libre creado se utiliza para reducir una cadena de cerca aceptores de electrones, que tienen mayor posteriormente redox potenciales. Estos pasos de transferencia de electrones son la fase inicial de una serie de reacciones de conversión de energía, en última instancia resulta en la producción de energía química durante la fotosíntesis .
La transformación de la energía luminosa en la separación de cargas
Centros de reacción están presentes en todos los verdes plantas , algas , y muchas bacterias . Aunque estas especies están separadas por miles de millones de años de evolución , los centros de reacción son homóloga para todas las especies fotosintéticas. En contraste, una gran variedad de Existen complejos captadores de luz entre las especies fotosintéticas. Las plantas verdes y las algas tienen dos tipos diferentes de centros de reacción que son parte de supercomplexes más grandes conocidos como fotosistema IP700 y fotosistema II P680. Las estructuras de estos supercomplexes son grandes, que implica múltiples complejos captadores de luz. El centro de reacción encuentra en Bacterias Rhodopseudomonas está actualmente entienden mejor, ya que fue el primer centro de reacción de estructura conocida y tiene menos cadenas polipeptídicas que los ejemplos de las plantas verdes.
Un centro de reacción se presenta en una forma tal que captura la energía de un fotón usando moléculas de pigmento y la convierte en una forma utilizable. Una vez que la energía de la luz ha sido absorbida directamente por las moléculas de pigmento, o que se les pasa por la transferencia de resonancia de un circundante complejo captador de luz, liberan dos electrones en un cadena de transporte de electrones.
La luz está compuesta de pequeños paquetes de energía llamados fotones . Si un fotón con la cantidad correcta de energía realiza un electrón, se elevará el electrón a un mayor nivel de energía. Los electrones son más estables en su nivel más bajo de energía, lo que también se llama su estado fundamental. En este estado, el electrón en la órbita es que tiene la menor cantidad de energía. Los electrones en los niveles de energía más altos pueden volver a estado fundamental de una manera análoga a una pelota que cae por una escalera. Al hacerlo, los electrones liberan energía. Este es el proceso que se explota por un centro de reacción fotosintética.
Cuando un electrón se eleva a un nivel superior de energía, disminución de la potencial de reducción de la molécula en la que reside el electrón se produce. Esto significa que la molécula tiene una mayor tendencia a donar electrones, la clave para la conversión de la energía luminosa en energía química. En las plantas verdes, la cadena de transporte de electrones que sigue tiene muchas aceptores de electrones incluidos feofitina, quinona, plastoquinona, bf citocromo, y ferredoxina, que dan lugar a la molécula reducida NADPH. El paso del electrón a través de la cadena de transporte de electrones también se traduce en el bombeo de protones (iones de hidrógeno) a partir de la cloroplasto de en el estroma lumen, resultando en un gradiente de protones a través de la membrana tilacoide que se puede utilizar para sintetizar ATP usando ATP sintasa. Tanto el ATP y NADPH se utilizan en el Ciclo de Calvin para fijar el dióxido de carbono en azúcares triosa.
Bacterias
Estructura
| Bacteriana centro de reacción fotosintética | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Bacteriana centro de reacción fotosintética | |||||||||
| Identificadores | |||||||||
| Símbolo | |||||||||
| Pfam | PF00124 | ||||||||
| InterPro | IPR000484 | ||||||||
| PROSITE | PDOC00217 | ||||||||
| SCOP | 1prc | ||||||||
| SUPERFAMILIA | 1prc | ||||||||
| TCDB | 3.e.2 | ||||||||
| |||||||||
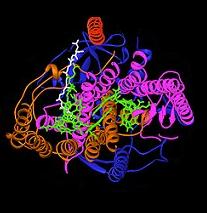
El centro de reacción fotosintético bacteriano ha sido un importante modelo para comprender la estructura y la química del proceso biológico de capturar energía de la luz. En la década de 1960, Roderick Clayton fue el primero para purificar el complejo de centro de reacción a partir de bacterias púrpuras. Sin embargo, la primera estructura cristalina se determinó en 1982 por Hartmut Michel, Johann Deisenhofer y Robert Huber para el que compartió el Premio Nobel en 1988. Esto también fue significativa, ya que era la primera estructura para cualquier complejo de proteínas de membrana.
Se encontraron cuatro subunidades diferentes que son importantes para la función del centro de reacción fotosintética. Las subunidades L y M, que se muestran en azul y púrpura en la imagen de la estructura, tanto abarcan el bicapa lipídica de la membrana plasmática. Ellos son estructuralmente similares entre sí, ambos con 5 transmembrana hélices alfa. Cuatro bacteriochlorophyll b (BChl-b) moléculas, dos B moléculas bacteriophaeophytin (HPB), dos moléculas quinonas (Q A y Q B), y un ion ferroso se asocian con la L y M subunidades. La subunidad H, que se muestra en oro, se encuentra en el lado citoplásmico de la membrana plasmática. Una subunidad citocromo, aquí no se muestra, contiene cuatro haems de tipo C y se encuentra en la superficie periplásmico (exterior) de la membrana. La sub-unidad de este último no es un motivo estructural general en las bacterias fotosintéticas. Las subunidades L y M se unen los cofactores funcionales y luminosos interactuar, mostradas aquí en verde.
Centros de reacción de diferentes especies bacterianas pueden contener alterado ligeramente bacterio-clorofila y cromóforos bacterio-feofitina como co-factores funcionales. Estas alteraciones causan cambios en el color de la luz que puede ser absorbida, creando así nichos específicos para la fotosíntesis. El centro de reacción contiene dos pigmentos que sirven para recoger y transferir la energía a partir de la absorción de fotones: BChl y Bph. BChl se asemeja más o menos la molécula de clorofila que se encuentra en las plantas verdes, pero, debido a las diferencias estructurales menores, su longitud de onda de absorción máxima se desplaza en el infrarrojos, con longitudes de onda tanto tiempo como 1000 nm. Bph tiene la misma estructura que BChl, pero el ion central de magnesio se sustituye por dos protones. Esta alteración provoca tanto un desplazamiento máximo de absorción y un potencial redox bajada.
Mecanismo

El proceso se inicia cuando la luz es absorbida por dos moléculas de BChl (una dímero) que se encuentran cerca de la lado periplásmico de la membrana. Este par de moléculas de clorofila, a menudo llamado el "par especial", absorbe fotones entre 870 nm y 960 nm, dependiendo de la especie y, por tanto, se denomina P870 (para las especies Rhodobacter sphaeroides) o P960 (por Rhodopseudomonas viridis), con P: la de "pigmento"). Una vez P absorbe un fotón, se expulsa un electrón, que se transfiere a través de otra molécula de Bchl a la BPh en la subunidad L. Esta inicial carga de separación produce una carga positiva en P y una carga negativa en la HPB. Este proceso se lleva a cabo en 10 picosegundos (10 -11 segundos).
Los cargos en el specialpair + y el BPh - podrían sufrir recombinación de carga en este estado. Esto perder el electrón de alta energía y convertir la energía de la luz absorbida en al calor . Varios factores de la estructura centro de reacción sirven para prevenir esto. En primer lugar, la transferencia de un electrón de BPh - para P960 + es relativamente lento en comparación con otros dos reacciones redox en el centro de reacción. Las reacciones más rápidas implican la transferencia de un electrón de BPh - (BPh - se oxida a BPh) a la quinona receptor de electrones (Q A), y la transferencia de un electrón a P960 + (P960 + se reduce a P960) de un grupo hemo en la subunidad citocromo encima del centro de reacción.
El electrón de alta energía que reside en la molécula de quinona fuertemente unido Q A se transfiere a una molécula de quinona intercambiable Q B. Esta molécula se asocia libremente con la proteína y es bastante fácil de separar. Dos de los electrones de alta energía se requieren para reducir totalmente Q B a QH 2, ocupando dos protones del citoplasma en el proceso. La quinona reducida QH 2 difunde a través de la membrana a otro complejo de proteína ( bc citocromo 1 -complex) donde se oxida. En el proceso el poder reductor de la QH 2 se utiliza para bombear protones a través de la membrana hacia el espacio periplásmico. Los electrones del citocromo bc 1 -complex se transfieren a continuación a través de un citocromo C soluble intermedia, llamada citocromo c 2, en el periplasma a la subunidad citocromo. Por lo tanto, el flujo de electrones en este sistema es cíclico.
Las plantas verdes
Fotosíntesis oxigénica
En 1772, el químico Joseph Priestley llevó a cabo una serie de experimentos relacionados con los gases implicados en la respiración y la combustión. En su primer experimento, encendió una vela y la colocó bajo un frasco de vuelta hacia arriba. Después de un corto período de tiempo, la vela quemada. Se llevó a cabo un experimento similar con un ratón en el espacio reducido de la vela encendida. Él encontró que el ratón murió poco tiempo después de la vela se había extinguido. Sin embargo, podría revivir el aire viciado por la colocación de las plantas verdes en la zona y exponiéndolos a la luz. Observaciones de Priestley, fueron algunos de los primeros experimentos que demostraron la actividad de un centro de reacción fotosintética.
En 1779, Jan Ingenhousz llevó a cabo más de 500 experimentos repartidos en 4 meses en un intento de entender lo que realmente estaba pasando. Él escribió a sus descubrimientos en un libro titulado Los experimentos sobre Vehículos. Ingenhousz tomó las plantas verdes y les sumerge en agua dentro de un tanque transparente. Se observó muchas burbujas que suben desde la superficie de las hojas cuando las plantas se expusieron a la luz. Ingenhousz recogió el gas que se desprende por las plantas y realizó varias pruebas diferentes en un intento de determinar cuál era el gas. La prueba que finalmente reveló la identidad del gas estaba colocando una conicidad latente en la muestra de gas y que tiene que volver a encender. Esta prueba demostró que era oxígeno, o, como Joseph Priestley había llamado, 'de- aire flogisticado '.
En 1932, el profesor Robert Emerson y un estudiante de pregrado, William Arnold, utilizaron una técnica de flash repetitivo para medir con precisión pequeñas cantidades de oxígeno desprendido por la clorofila en la Chlorella algas. Su experimento demostró la existencia de una unidad fotosintética. Gaffron y Wohl tarde interpretaron el experimento y se dieron cuenta de que la luz absorbida por el aparato fotosintético fue trasladado. Esta reacción se produce en el centro de reacción del fotosistema II y tiene lugar en las cianobacterias, algas y plantas verdes.
Fotosistema II

Fotosistema II es el fotosistema que genera los dos electrones que eventualmente reducir NADP + en Ferredoxina-NADP-reduktase. Fotosistema II está presente en las membranas tilacoides dentro de los cloroplastos, el sitio de la fotosíntesis en las plantas verdes. La estructura del fotosistema II es notablemente similar a la del centro de reacción bacteriana, y se teoriza que comparten un ancestro común.
El núcleo del fotosistema II consiste en dos subunidades denominadas D1 y D2. Estas dos subunidades son similares a la L y M subunidades presentes en el centro de reacción bacteriana. Fotosistema II difiere del centro de reacción bacteriana en que tiene muchas subunidades adicionales que se unen clorofilas adicionales para aumentar la eficiencia. La reacción global catalizada por el fotosistema II es:
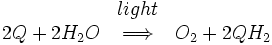
Q representa plastoquinona, la forma oxidada de Q. QH 2 representa plastoquinol, la forma reducida de Q. Este proceso de reducción de quinona es comparable a la que tiene lugar en el centro de reacción bacteriana. Fotosistema II obtiene electrones por el agua oxidante en un proceso llamado fotólisis. El oxígeno molecular es un subproducto de este proceso, y es esta reacción que suministra el atmósfera con oxígeno. El hecho de que el oxígeno de las plantas verdes se originó a partir de agua se dedujo primero por el bioquímico estadounidense de origen canadiense Martin David Kamen. Él utilizó una natural, isótopos estables de oxígeno, O 18 a trazar el camino del oxígeno, a partir de agua al oxígeno molecular gaseoso. Esta reacción está catalizada por un centro reactivo en el fotosistema II que contiene cuatro de manganeso iones .
La reacción comienza con la excitación de un par de moléculas de clorofila similares a aquellos en el centro de reacción bacteriana. Debido a la presencia de clorofila a, en lugar de bacteriochlorophyll, fotosistema II absorbe luz a una longitud de onda más corta. El par de moléculas de clorofila en el centro de reacción se refiere a menudo como P680. Cuando el fotón ha sido absorbido, el electrón de alta energía resultante se transfiere a una molécula feofitina cerca. Esto está por encima y a la derecha de la pareja en el diagrama y es de color gris. El electrón se desplaza desde la molécula feofitina través de dos moléculas plastoquinona, el primer fuertemente unido, el segundo débilmente unida. La molécula fuertemente unido se muestra por encima de la molécula feofitina y es de color rojo. La molécula es unido débilmente a la izquierda de esto y también es de color rojo. Este flujo de electrones es similar a la del centro de reacción bacteriana. Se requieren dos electrones para reducir completamente la molécula plastoquinona débilmente unida a QH 2, así como la absorción de dos protones.
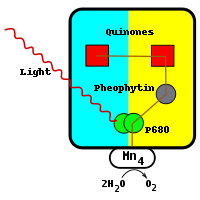
La diferencia entre el fotosistema II y el centro de reacción bacteriana es la fuente del electrón que neutraliza el par de moléculas de clorofila a. En el centro de reacción bacteriana, el electrón se obtiene a partir de un grupo hemo compuesto reducido en una subunidad del citocromo o de una proteína soluble en agua citocromo-c.
Una vez separación de carga fotoinducida ha tenido lugar, la molécula P680 lleva un positivo cargo . P680 es un muy fuerte oxidantes y extractos electrones de dos de agua moléculas que se unen en el centro de manganeso directamente debajo del par. Este centro, por debajo y a la izquierda de la pareja en el diagrama, contiene cuatro iones de manganeso, un calcio iónico, un cloruro de iones, y una residuo de tirosina. El manganeso es eficaz porque es capaz de existir en cuatro estados de oxidación: Mn 2+, 3+ Mn, Mn y Mn 4+ 5+. El manganeso también forma enlaces fuertes con las moléculas que contienen oxígeno como el agua.
Cada vez que el P680 absorbe un fotón, que emite un electrón, ganando una carga positiva. Esta carga se neutraliza por la extracción de un electrón desde el centro de manganeso, que se encuentra directamente debajo de él. El proceso de oxidación de dos moléculas de agua requiere cuatro electrones. Las moléculas de agua que se oxidan en el centro de manganeso son la fuente de los electrones que reduzcan las dos moléculas de Q para QH 2. Hasta la fecha, este centro catalítico de la disociación del agua no puede ser reproducido por cualquier catalizador por el hombre.
Fotosistema I
Después de que el electrón ha dejado fotosistema II se transfiere a un citocromo complejo b6f y luego a plastocianina, de un azul de cobre y la proteína portadora de electrones. El complejo plastocianina lleva el electrón que neutralizará el par en el próximo centro de reacción, fotosistema I.
Al igual que con el fotosistema II y el centro de reacción bacteriana, un par de moléculas de clorofila inicia una separación de carga fotoinducida. Este par se conoce como P700. 700 es una referencia a la longitud de onda a la que las moléculas de clorofila absorben la luz al máximo. El P700 se encuentra en el centro de la proteína. Una vez separación de carga fotoinducida se ha iniciado, el electrón viaja por un camino a través de una molécula de clorofila α situado directamente encima de la P700, a través de una molécula de quinona situado directamente encima de que, a través de tres grupos 4Fe-4s, y, finalmente, a un complejo de ferredoxina intercambiables. Ferredoxina es una proteína soluble que contiene un clúster de 2Fe-2S coordinado por cuatro residuos de cisteína. La carga positiva a la izquierda en el P700 se neutraliza por la transferencia de un electrón de plastocianina. Así, la reacción general catalizada por el fotosistema I es:
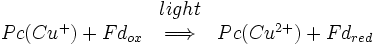
La cooperación entre los fotosistemas I y II crea un flujo de electrones entre H 2 O a NADP +. Esta vía se llama 'Z-sistema »porque el diagrama redox de P680 a P700 se asemeja a la letra z.