

Rétine
Contexte des écoles Wikipédia
Ce contenu de Wikipedia a été sélectionné par SOS Enfants d'aptitude dans les écoles à travers le monde. Mères SOS chaque regard après une une famille d'enfants parrainés .
| Rétine | |
|---|---|
 | |
| Droit oeil humain de vue en coupe. Courtoisie NIH National Eye Institute. De nombreux animaux ont des yeux différents de l'oeil humain. | |
| Gray | objet # 225 1014 |
| Artère | artère centrale de la rétine |
| MeSH | Rétine |
| Dorlands / Elsevier | Rétine |
- Retina peut également se référer à la Kodak Retina et Apple Retina Display.
La rétine de vertébrés (pron .: / r ɛ t ɪ n ə / RET -nuh, pl. rétines, pron .: / r ɛ t Je n Je /; de Rete latin, signifiant «net») est une couche sensible à la lumière de tissu, en alignant la surface intérieure de l' oeil . L'optique de l'oeil de créer une image du monde visuel sur la rétine, qui sert la même fonction que le film dans une caméra. Suppression de la rétine lumière déclenche une cascade d'événements chimiques et électriques qui déclenchent finalement l'influx nerveux. Ceux-ci sont envoyés à différents centres visuels du cerveau à travers les fibres de la nerf optique.
Chez les vertébrés le développement embryonnaire, la rétine et le nerf optique proviennent comme des excroissances de l'élaboration de cerveau , de sorte que la rétine est considérée comme faisant partie de la système nerveux central (SNC) et est en fait le tissu cérébral. Ce est la seule partie du système nerveux central qui peut être visualisée de manière non invasive.
La rétine est une structure stratifiée de plusieurs couches de neurones interconnectés par des synapses . Les seuls neurones qui sont directement sensibles à la lumière sont les les cellules photoréceptrices. Il se agit principalement de deux types: le tiges et cônes. Tiges fonctionnent surtout en faible lumière et offrent une vision en noir et blanc, tandis que les cônes soutiennent la vision diurne et la perception des couleurs. Un troisième type de photorécepteur beaucoup plus rare, le cellules ganglionnaires photosensible, est important pour les réponses réflexes à la lumière du jour.
Des signaux neuronaux des cônes et des bâtonnets subissent un traitement par d'autres neurones de la rétine. La sortie prend la forme de potentiels d'action en les cellules ganglionnaires de la rétine dont axones forment le nerf optique. Plusieurs caractéristiques importantes de perception visuelle peut être attribuée à l'encodage de la rétine et le traitement de la lumière.
Anatomie de la rétine des vertébrés

La rétine des vertébrés a dix couches distinctes. Du plus proche au plus éloigné du corps vitré - ce est, de la plus proche à l'extérieur avant de la tête vers l'intérieur et à l'arrière de la tête:
- Membrane basale élaboré par - limitant membrane interne Les cellules de Müller
- Nerve couche de fibres - axones des noyaux des cellules ganglionnaires (à noter qu'il existe une mince couche de palettes de cellules Müller entre cette couche et la membrane limitante interne)
- couche de cellules ganglionnaires - contient des noyaux des cellules ganglionnaires, les axones qui deviennent les fibres du nerf optique pour les messages et certaines cellules amacrines déplacées
- Couche plexiforme interne - contient la synapse entre les axones des cellules bipolaires et les dendrites des cellules amacrines et ganglionnaires.
- Couche nucléaire interne - contient les noyaux et les corps cellulaires environnantes (péricaryons) de la cellules bipolaires.
- Couche plexiforme externe - projections de bâtonnets et les cônes se terminant dans le sphérules de cônes et bâtonnets pédicule, respectivement. Celles-ci forment des synapses avec les dendrites des cellules bipolaires. Dans le région maculaire, ceci est connu comme la couche de fibres de Henle.
- Couche nucléaire extérieure - corps cellulaires des bâtonnets et des cônes
- Membrane limitante externe - couche qui sépare les parties de segments internes des photorécepteurs de leur noyau de la cellule
- couche des photorécepteurs - tiges / cônes
- Épithélium pigmentaire rétinien - une seule couche de cellules cubiques (avec extrusions non représenté sur le schéma)
Ceux-ci peuvent être simplifiées en 4 grandes étapes de traitement: photoréception, transmission cellules bipolaires, la transmission aux cellules ganglionnaires qui contiennent également des photorécepteurs, les cellules ganglionnaires photosensibles et la transmission le long du nerf optique. A chaque étape synaptique il ya aussi communicantes latéralement horizontal et cellules amacrines.
Le nerf optique est un appareil central de nombreux axones des cellules ganglionnaires reliant principalement à la Corps genouillé latéral, une station de relais visuel dans le diencéphale (l'arrière du cerveau antérieur). Il projette également à la colliculus supérieur, le noyau suprachiasmatique, et la noyau du faisceau optique. Il passe à travers les autres couches qui créent la Disque optique chez les primates.
Structures supplémentaires, non directement liés à la vision, se retrouvent comme des excroissances de la rétine dans certains groupes de vertébrés. Dans les oiseaux , le Pecten est une structure vasculaire de forme complexe qui fait saillie de la rétine dans le vitré; il fournit de l'oxygène et des nutriments à l'œil, et peut également aider à la vision. Reptiles ont une structure similaire, mais beaucoup plus simple,.
Un mnémotechnique pour se rappeler les couches de la rétine:
| M y | Les cellules de Müller |
| N Erves | Les fibres nerveuses |
| G et | Ganglions |
| Je n | Plexiforme interne |
| K n ots | Nucléaire interne |
| O utside | Plexiforme externe |
| N otre | Nucléaire externe |
| E asy | Membrane limitante externe |
| PRATIQUE | Photorécepteurs |
| R evue | Épithélium pigmentaire rétinien |
Structure physique de la rétine humaine
Chez l'homme adulte, la rétine entière est d'environ 72% environ d'une sphère de 22 mm de diamètre. La rétine entière contient environ 7 millions de cônes et 75-150000000 tiges. Le disque optique, une partie de la rétine appelé parfois "l'angle mort" parce qu'il manque de photorécepteurs, est situé à la papille optique, une zone nasale où les fibres du nerf optique quittent l'œil. Il apparaît comme une zone blanche ovale de 3mm². Temporelle (dans le sens des temples) pour ce disque est le macula. En son centre est le fovéa, une fosse qui est responsable de notre vision centrale, mais est en fait moins sensibles à la lumière en raison de son manque de tiges. Humain et non humain primates possèdent une fovéa par opposition à certaines espèces d'oiseaux comme les faucons qui sont réellement bifoviate et les chiens et les chats qui ne possèdent pas de fovéa, mais une bande centrale connue sous le nom série visuelle. Autour de la fovéa étend la rétine centrale pour environ 6 mm et ensuite la rétine périphérique. Le bord de la rétine est définie par la serrata ora. La longueur de l'un à l'autre ora (ou macula), la zone la plus sensible le long de l'horizontale méridienne est d'environ 32 mm.

Dans la section de la rétine est pas supérieure à 0,5 mm d'épaisseur. Il dispose de trois couches de les cellules nerveuses et de deux synapses, y compris l'unique, synapses de ruban. Le nerf optique transmet la cellules ganglionnaires axones dans le cerveau et les vaisseaux sanguins qui se ouvrent dans la rétine. Les cellules ganglionnaires se trouvent plus à l'intérieur de la rétine, tandis que les cellules photoréceptrices se situent plus à l'extérieur. En raison de cet agencement de contre-intuitif, la lumière doit d'abord passer à travers et autour des cellules ganglionnaires et à travers l'épaisseur de la rétine, (y compris ses vaisseaux capillaires, non représenté) avant d'atteindre les bâtonnets et les cônes. Toutefois, il ne passe pas par la ou l'épithélium choroïde (qui sont tous deux opaque).
Le des globules blancs dans le capillaires devant les photorécepteurs peuvent être perçus comme de minuscules points lumineux en mouvement quand on regarde dans la lumière bleue. Ceci est connu comme la champ bleu phénomène entoptique (ou le phénomène de Scheerer).
Entre la couche des cellules ganglionnaires et les bâtonnets et les cônes, il existe deux couches de neuropils où les contacts synaptiques sont faites. Les couches sont le neuropile et la couche plexiforme externe couche interne plexiforme. Dans l'extérieur des bâtonnets et des cônes se connecter à des cellules bipolaires se étendant verticalement, et les cellules horizontales orientées horizontalement connecter à cellules ganglionnaires.
La rétine centrale est conique dominé et la rétine périphérique est dominé par tige. Au total, il ya environ sept millions de cônes et une centaine de millions de bâtonnets. Au centre de la macula est la fosse fovéale où les cônes sont la plus petite et en une mosaïque hexagonale, la plus efficace et la plus forte densité. Sous la fosse les autres couches de la rétine sont déplacées, avant de construire le long de la pente de la fovéa jusqu'à la jante de la fovéa ou parafovéa qui est la partie la plus épaisse de la rétine. La macula a une pigmentation jaune de pigments de dépistage et est connue comme la macula lutea. La zone entourant directement la fovéa a la plus forte densité de tiges convergeant sur bipolaires simples. Depuis les cônes ont une puissance bien moindre de la fusion de signaux, la fovéa permet la vision la plus nette l'œil peut atteindre.
Bien que la tige et les cônes sont une mosaïque de toutes sortes, la transmission des récepteurs à bipolaires aux cellules ganglionnaires ne est pas direct. Depuis, il ya environ 150 millions de récepteurs et seulement 1 million de fibres du nerf optique, il doit y avoir convergence et mélangeant ainsi des signaux. En outre, l'action horizontale des cellules amacrines et horizontales peut permettre à une zone de la rétine pour commander un autre (par exemple une inhibition de stimulus autre). Cette inhibition est essentiel à la somme des messages envoyés vers les régions supérieures du cerveau. Dans certains vertébrés inférieurs, (par exemple, le pigeonnier ) il ya un contrôle «centrifuge» de messages - ce est, une couche peut contrôler une autre, ou des régions supérieures du cerveau peut conduire les cellules nerveuses de la rétine, mais chez les primates cela ne se produit pas.
Vertébrés et céphalopodes différences de la rétine
La rétine des vertébrés est inversée en ce sens que les cellules de détection de lumière se asseoir à l'arrière de la rétine, de sorte que la lumière doit traverser des couches de neurones et capillaires avant d'atteindre les bâtonnets et les cônes. En revanche, la céphalopode rétine a les photorécepteurs sur le côté avant de la rétine, avec traitement de neurones et capillaires derrière eux. Pour cette raison, céphalopodes ne ont pas de angle mort.
La rétine de céphalopodes ne provient pas comme une excroissance du cerveau, comme celui des vertébrés fait. On peut soutenir que cette différence montre que vertébrés et céphalopodes yeux ne sont pas homologue mais ont évolué séparément.
En 2009 Kröger anatomique a montré dans Zebrafish que si l'entente est non adaptatif inversé en ce qu'il crée évitable diffusion de la lumière (et donc la perte de lumière et de flou de l'image), il a des avantages d'économie d'espace pour les animaux de petite yeux dans lesquels il existe un corps vitré minime, car l'espace entre la lentille et segments extérieurs sensibles à la lumière des photorécepteurs est complètement rempli avec des cellules rétiniennes.
La différence entre les vertébrés et les céphalopodes rétines présente un puzzle intéressant de chemin évolutif qui ne est pas encore totalement réglée. Dans une perspective évolutionniste, une structure alambiquée comme la rétine inversée peuvent généralement se produire à la suite de deux procédés alternatifs; (A) un "bon" compromis avantageux entre concurrents limitations fonctionnelles, ou (B) comme une relique historique de inadapté le chemin tortueux de l'évolution d'organes et de transformation. Vision est une adaptation importante chez les vertébrés supérieurs. Par conséquent, si la rétine est en effet "câblé tort" ou "mal conçu" (d'un point d'ingénierie de vue optique), il est sensé de regarder pour qu'il y ait éventuellement un avantage physiologique très important. Une telle suggestion est fondée sur l'argument que le processus d'amplification photorécepteur mammifère nécessite de grandes quantités d'énergie métabolique, et par conséquent, il nécessite apport massif et homogène de sang. En effet, un réseau unique de vaisseaux sanguins est bien adapté pour fournir la couche de photorécepteur avec de grandes quantités de sang. Cela montre que la rétine inversée est une adaptation pour fournir des quantités abondantes de l'oxygène à la rétine correspondant à ses exigences élevées en énergie et avec un bon maintien du pigment rétinien épithélium (RPE), les cellules contre les dommages photo-oxydant, ce qui, tandis que sur la face de il est exacerbé par le sang riche en oxygène dans la choroïde, ne est-le-moins éliminé par le processus de recyclage de disque opsin il permet. Ce dernier effet permet aux cellules photoréceptrices d'avoir une longue (c. décennies) de durée de vie utile. Les céphalopodes ont une rétine non inversé qui est comparable dans la résolution de la puissance aux yeux de nombreux vertébrés, cependant, les photorécepteurs sont pas maintenu, et cela oblige tous les invertébrés soit avoir une vie courte (de quelques années) dans un environnement photopique ou passer la plupart de leur vie dans l'obscurité. Une troisième possibilité, d'avoir tige-yeux facilement remplacés (certains homards) ou rétines (certaines araignées, comme Deinopis) est rare.
Physiologie
Une image est produite par l'excitation des motifs cônes et bâtonnets de la rétine. L'excitation est générée par le système neuronal et les différentes parties du cerveau travaillant en parallèle pour former une représentation de l'environnement extérieur dans le cerveau.
Les cônes réagissent à la lumière et la médiation haute résolution vision des couleurs lors de la lumière du jour (aussi appelés vision photopique). Les tiges sont saturés au niveau de lumière du jour et ne contribuent pas à la vision du motif. Cependant, les tiges ne répondent pour atténuer la lumière et la médiation basse résolution, la vision monochromatique sous très faibles niveaux d'éclairage (appelés vision scotopique). L'éclairage dans la plupart des paramètres de bureau se situe entre ces deux niveaux et est appelé vision mésopique. A ces niveaux de lumière, les deux cônes et les bâtonnets contribuent activement des informations de motif à cette sortie de l'œil. Quelle contribution les informations de tige fait à la vision de modèle dans ces circonstances ne est pas claire.
La réponse des cônes à différentes longueurs d'onde de la lumière est appelé leur sensibilité spectrale. Dans la vision humaine normale, la sensibilité spectrale d'un cône tombe dans l'un des trois sous-groupes. On les appelle souvent bleu, vert, et les cônes rouges, mais avec plus de précision sont à court, moyen et long longueur d'onde sous-groupes de cônes sensibles. Ce est un manque d'un ou plusieurs des sous-types de cônes qui provoque les individus à avoir des lacunes dans la vision des couleurs ou différents types de daltonisme . Ces personnes ne sont pas aveugles aux objets d'une couleur particulière, mais l'expérience de l'incapacité à distinguer entre deux groupes de couleurs qui peuvent être distingués par des gens ayant une vision normale. Les humains ont trois types de cônes différents ( une vision trichromatique), tandis que la plupart des autres mammifères manquent cônes avec pigment sensible au rouge et ne ont donc plus pauvres (vision dichromatique) de couleur. Cependant, certains animaux ont quatre sous-groupes spectrales, par exemple la truite ajoute un sous-groupe de l'ultraviolet à court, moyen et long terme sous-groupes qui sont similaires aux humains. Certains poissons sont sensibles à la polarisation de la lumière ainsi.
Lorsque la lumière tombe sur un récepteur, il envoie une réponse proportionnelle à synaptiquement cellules bipolaires qui à son tour signalent les cellules ganglionnaires de la rétine. Les récepteurs sont aussi «réticulé» par cellules horizontales et les cellules amacrines, qui modifient le signal synaptique avant que les cellules ganglionnaires. signaux de bâtonnets et des cônes sont entremêlées et se combinent, bien que les tiges sont surtout actifs dans des conditions très mal éclairées et saturer en plein jour, alors que les cônes fonctionnent dans l'éclairage plus lumineux parce qu'ils ne sont pas assez sensibles pour travailler à des niveaux de très faible luminosité.
En dépit du fait que tous sont des cellules nerveuses, seules les cellules ganglionnaires de la rétine et quelques cellules amacrines créer des potentiels d'action. Dans les photorécepteurs, exposition à la lumière hyperpolarise la membrane dans une série de glissements classés. Le segment externe de la cellule contient une photopigment. L'intérieur de la cellule, les niveaux normaux de guanosine monophosphate cyclique (GMPc) garder le canal Na + ouvert et donc dans l'état de repos de la cellule est dépolarisée. Le photon provoque le rétinien lié à la protéine récepteur à isomériser à trans-rétinal. Cela provoque récepteur pour activer multiple G-protéines. Cela provoque à son tour la sous-unité Ga de la protéine à activer une phosphodiesterase (PDE6), ce qui dégrade le GMPc, ce qui entraîne la fermeture de Na + chaînes de nucléotides cycliques-dépendants ion (GCN). Ainsi, la cellule est hyperpolarisé. La quantité de neurotransmetteur libéré est réduit dans la lumière et augmente brillant comme niveau de luminosité diminue. Le photopigment réelle est blanchie loin dans la lumière vive et seulement remplacé comme un processus chimique, donc dans une transition de la lumière vive à l'obscurité l'œil peut prendre jusqu'à 30 minutes pour atteindre la sensibilité complète (voir Adaptation (oeil)).
Dans les cellules ganglionnaires de la rétine, il existe deux types de réponses, en fonction de la champ récepteur de la cellule. Les champs récepteurs des cellules ganglionnaires de la rétine comporter une zone approximativement circulaire centrale, où la lumière a un effet sur la mise à feu de la cellule, et une ambiance annulaire, où la lumière a l'effet inverse sur la mise à feu de la cellule. Dans les cellules ON, une augmentation de l'intensité lumineuse au centre du champ récepteur provoque le taux d'allumage augmente. Dans les cellules OFF, elle rend diminuer. Dans un modèle linéaire, ce profil de réponse est bien décrite par une Différence gaussienne et est la base de Les algorithmes de détection de bord. Au-delà de cette simple différence cellules ganglionnaires se différencient également par la sensibilité chromatique et du type de sommation spatiale. Cellules présentant sommation spatiale linéaire sont appelées cellules X (également appelé parvocellulaire, P, ou des cellules midget ganglionnaires), et ceux montrant sommation non linéaire sont des cellules Y (également appelé magnocellulaire, M, ou des cellules ganglionnaires de parasol de la rétine), bien que la correspondance entre X et Y (cellules de la rétine de chat) et des cellules P et M (dans la rétine des primates) ne est pas aussi simple que cela semblait une fois.
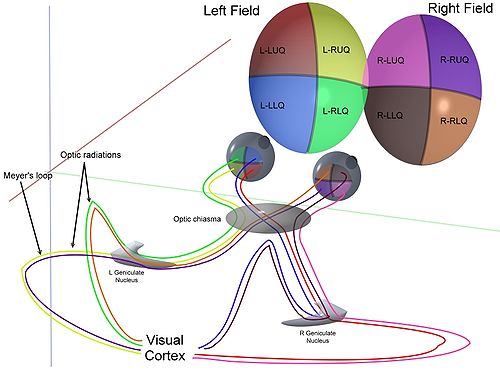
Dans le transfert de signaux visuels au cerveau, la voie visuelle, la rétine est verticalement divisée en deux, une temporel (plus proche de la tempe) et une moitié nasale (plus près du nez) moitié. Les axones traversent nasale de la moitié du cerveau à la chiasma optique de se joindre à axones de la moitié temporelle de l'autre œil avant de passer dans le corps genouillé latéral.
Bien qu'il existe plus de 130 millions de récepteurs de la rétine, il ya seulement environ 1,2 millions de fibres (axones) dans le nerf optique; une grande quantité de pré-traitement est effectué à l'intérieur de la rétine. La fovéa produit l'information la plus précise. Malgré occupant environ 0,01% du champ visuel (moins de 2 ° de angle visuel), environ 10% des axones dans le nerf optique sont consacrées à la fovéa. La limite de résolution de la fovéa a été déterminée à environ 10 000 points. La capacité de l'information est estimé à 500 000 bits par seconde (pour plus d'informations sur les bits, voir théorie de l'information), sans couleur ou autour de 600 000 bits par seconde, y compris la couleur.
Le codage spatial

La rétine ne se contente pas envoyer une image vers le cerveau. La rétine code spatialement (compresse) l'image pour se adapter à la capacité limitée du nerf optique. Compression est nécessaire parce qu'il ya 100 fois plus cellules photoréceptrices que les cellules ganglionnaires comme mentionné ci-dessus. La rétine fait en "Décorrélation" les images entrantes dans une manière qui sera décrite ci-dessous. Ces opérations sont effectuées par les structures surround centre Mise en œuvre par les cellules bipolaires et ganglionnaires.
Il existe deux types de structures surround centre dans les centres off-rétine-sur-centres et. Sur-centres ont un centre positivement pondérée et un Surround pondérée négativement. Off-centres sont tout le contraire. Pondération positive est plus communément connu comme excitateur et la pondération négative est plus communément connu sous le nom d'inhibition .
Ces structures centre surround ne sont pas physiques dans le sens où on ne peut pas les voir par coloration des échantillons de tissus et d'examiner l'anatomie de la rétine. Les structures centre surround sont logiques (ce est à dire, mathématiquement résumé) dans le sens où ils dépendent des forces de connexion entre ganglion et les cellules bipolaires. On pense que les forces de connexion entre les cellules est provoquée par le nombre et les types de les canaux ioniques noyés dans le synapses entre les cellules ganglionnaires et bipolaires. Voir Champ récepteur pour les chiffres et plus d'informations sur les structures surround centre.
Les structures centre surround sont mathématiquement équivalente à la algorithmes de détection de pointe utilisées par les programmeurs informatiques pour extraire ou améliorer les bords dans une photographie numérique. Ainsi la rétine effectue des opérations sur l'image pour améliorer les bords des objets dans son champ visuel. Par exemple, dans une image d'un chien, un chat et une voiture, ce sont les bords de ces objets qui contiennent le plus d'informations. Pour que les fonctions plus élevées dans le cerveau (ou dans un ordinateur, d'ailleurs) pour extraire et classer des objets tels qu'un chien et un chat, la rétine est la première étape pour séparer les différents objets dans la scène.
A titre d'exemple, ce qui suit matrice est au cœur de l'ordinateur algorithme qui met en oeuvre la détection des contours. Cette matrice est équivalente à l'ordinateur central de la structure d'ambiance. Dans cet exemple, chaque boîte (élément) dans cette matrice serait reliée à une photorécepteur. Photorécepteur dans le centre du récepteur est en cours de traitement. Le centre photorécepteur est multiplié par le facteur 1 de poids. Les photorécepteurs environnantes sont les «plus proches voisins» au centre et sont multipliés par la valeur -1/8. La somme de tous neuf de ces éléments est finalement calculée. Cette sommation est répétée pour chaque photorécepteur dans l'image en décalant la gauche vers la fin d'une ligne et ensuite vers le bas pour la ligne suivante.
| -1/8 | -1/8 | -1/8 |
| -1/8 | 1 | -1/8 |
| -1/8 | -1/8 | -1/8 |
La somme totale de cette matrice est zéro si toutes les entrées en provenance des photorécepteurs neuf sont la même valeur. Le résultat de zéro indique que l'image était uniforme (non-changement) dans ce petit patch. Sommes négatives ou positives signifient quelque chose variait (changement) dans ce petit lopin de neuf photorécepteurs.
La matrice ci-dessus ne est qu'une approximation de ce qui se passe vraiment à l'intérieur de la rétine. Les différences sont:
- L'exemple ci-dessus est appelé «équilibré». Le terme pondérées signifie que la somme des poids négatifs est égal à la somme des poids positifs de sorte qu'ils se annulent parfaitement. Les cellules ganglionnaires de la rétine sont presque jamais parfaitement équilibré.
- Le tableau est carré tandis que les structures centre surround dans la rétine sont circulaires.
- Les neurones fonctionnent sur trains de potentiels en descendant cellule nerveuse axones. Les ordinateurs fonctionnent sur un seul Nombre à virgule flottante qui est essentiellement constante de chaque entrée pixel. (Le pixel de l'ordinateur est essentiellement l'équivalent d'un photorécepteur biologique).
- La rétine effectue tous ces calculs en parallèle pendant que l'ordinateur fonctionne sur chaque pixel une à la fois. Il n'y a pas de sommations répétées et le déplacement car il y aurait dans un ordinateur.
- Enfin, la horizontal et cellules amacrines jouent un rôle important dans ce processus, mais qui ne est pas représenté ici.
Voici un exemple d'une image d'entrée et comment détection de bord se modifier.
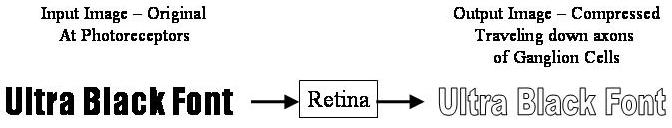
Une fois que l'image est codé spatialement par les structures centre surround, le signal est envoyé sur le nerf optique (par l'intermédiaire des axones des cellules ganglionnaires) à travers le chiasma à l'LGN ( noyau genouillé latéral). La fonction exacte du LGN est inconnu pour le moment. La sortie du LGN est ensuite envoyé à l'arrière du cerveau. Plus précisément la sortie de la CGL "rayonne" hors de la V1 Cortex visuel primaire.
Simplifié Flux de signal: photorécepteurs → → bipolaire ganglion → → Chiasm LGN → V1 cortex
Maladies et troubles
Il ya beaucoup de maladies ou de troubles héréditaires et acquis qui peuvent affecter la rétine. Certains d'entre eux comprennent:
- La rétinite pigmentaire est un groupe de maladies génétiques qui affectent la rétine et provoque la perte de la vision nocturne et la vision périphérique.
- La dégénérescence maculaire décrit un groupe de maladies caractérisées par une perte de la vision centrale en raison de la mort ou de la réduction des cellules dans le macula.
- Dystrophie cône tige (CORD) décrit un certain nombre de maladies où la perte de vision est provoquée par une détérioration de la cônes et / ou des tiges dans la rétine.
- En séparation de la rétine, la rétine se détache de l'arrière du globe oculaire. Ignipuncture est une méthode de traitement pas à jour. Le terme décollement de la rétine est utilisé pour décrire une séparation de la rétine neurosensorielle de la épithélium pigmentaire rétinien. Il existe plusieurs méthodes de traitement modernes de fixation d'un décollement de la rétine: rétinopexie pneumatique, indentation, cryothérapie, photocoagulation au laser et pars plana vitrectomie.
- Tant l'hypertension et le diabète sucré peut provoquer des dommages aux petits vaisseaux sanguins qui alimentent la rétine, conduisant à rétinopathie hypertensive et la rétinopathie diabétique.
- Rétinoblastome est un cancer de la rétine.
- Maladies de la rétine à chiens comprennent dysplasie de la rétine, atrophie rétinienne progressive, et soudain acquis une dégénérescence rétinienne.
- Lipémie rétinienne est un aspect blanc de la rétine, et peut se produire par dépôt lipidique en déficience en lipoprotéine lipase.
- La rétinopathie diabétique
Diagnostic et traitement
Un certain nombre d'instruments différents sont disponibles pour le diagnostic des maladies et troubles affectant la rétine. Ophtalmoscopie et la photographie de fundus sont utilisés pour examiner la rétine. Récemment, optique adaptative a été utilisé pour l'image bâtonnets et les cônes individuels dans la rétine humaine vivante et une société basée en Ecosse ont conçu une technologie qui permet aux médecins d'observer la rétine complète sans aucune gêne pour les patients.
Le électrorétinogramme est utilisé pour mesurer Les activités de la rétine non invasive électrique, qui est affectée par certaines maladies. Une technologie relativement nouvelle, devient maintenant largement disponible, est tomographie par cohérence optique (OCT). Cette technique non invasive permet d'obtenir un 3D volumétrique ou la résolution haute transversale tomogramme de la structure fine de la rétine avec histologique qualité.

Le traitement dépend de la nature de la maladie ou du trouble. La transplantation de rétines a été tenté, mais sans grand succès. À MIT, l'Université de Californie du Sud, l'Université RWTH Aachen, et de la Université de Nouvelle-Galles du Sud, une «rétine artificielle" est en cours de développement: un implant qui contournera les photorécepteurs de la rétine et de stimuler directement les cellules nerveuses attachés, avec des signaux à partir d'un appareil photo numérique.
Approvisionnement sanguin rétinien
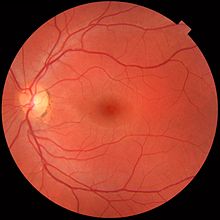
Il existe deux circulations, les deux fournies par l'artère ophtalmique. La circulation de l'uvée compose des artères entrer dans le monde en dehors du nerf optique, ceux-ci fournissent l'uvée et des couches extérieures et intermédiaires de la rétine. La circulation rétinienne, d'autre part, fournit la couche intérieure de la rétine et passe avec le nerf optique en tant que branche de l'artère ophtalmique appelé l'artère centrale de la rétine. La structure unique de la les vaisseaux sanguins de la rétine a été utilisé pour identification biométrique.
La géométrie topographique vasculaire dans la rétine est connu pour se conformer aux principes structurels qui sont liés à certaines propriétés physiques. L'analyse de la structure géométrique est très important que les écarts par rapport aux principes optimales peuvent indiquer certaines maladies cardiovasculaires, comme l'hypertension et l'athérosclérose; une analyse complète est donnée par Patton et al. (2006). L'identification des bifurcations vasculaires est l'une des étapes de base à cette analyse. Azzopardi et Petkov (2011) proposent une ordinateur algorithme de vision qui détecte automatiquement ces caractéristiques rétiniennes. Leurs résultats sont évalués contre le les données de réalité de terrain de bifurcations vasculaires d'images du fond d'œil de la rétine qui sont obtenus à partir de la paramètres d'entraînement.
Recherche
George Wald, Haldan Keffer Hartline et Ragnar Granit a remporté le 1967 Prix Nobel de physiologie ou médecine pour leur recherche scientifique sur la rétine.
Une récente Université de Pennsylvanie étude a calculé le approximative bande passante de rétines humaines est 8,75 mégabits par seconde, alors que Guinée porcs rétines transfert à 875 kilobits.
MacLaren & Pearson et ses collègues University College London et Moorfields Eye Hospital à Londres a montré en 2006 que cellules photoréceptrices pourraient être transplantés avec succès dans la rétine de souris si les cellules du donneur sont à un stade critique de développement. Récemment Ader et ses collègues ont montré à Dublin utilisant le microscope électronique que les photorécepteurs greffés formés connexions synaptiques.
En 2012 Sebastian Seung et son laboratoire à MIT ont lancé EyeWire, une ligne Citoyen jeu de la science où les joueurs tracent neurones dans la rétine. Les objectifs du projet sont EyeWire pour identifier les types de cellules spécifiques dans les grandes classes connues de cellules rétiniennes, et cartographier les connexions entre les neurones de la rétine, ce qui permettra de déterminer comment la vision fonctionne.
La thérapie génique de la rétine
La thérapie génique prometteuse comme une avenue potentiel de guérir un large éventail de maladies de la rétine. Cela implique l'utilisation d'un virus non infectieux de la navette d'un gène dans une partie de la rétine. Recombinant virus (rAAV) des vecteurs adéno-associés possèdent un certain nombre de caractéristiques qui les rendent idéales pour la thérapie génique de la rétine, y compris une absence de pathogénicité, l'immunogénicité minimale, et la capacité de transduire des cellules post-mitotiques de manière stable et efficace. des vecteurs rAAV sont de plus en plus utilisées pour leur capacité à médier la transduction efficace de épithélium pigmentaire rétinien (EPR), cellules photoréceptrices et les cellules ganglionnaires de la rétine. Chaque type de cellule peut être spécifiquement ciblé par le choix de la combinaison appropriée d'AAV sérotype, promoteur et site d'injection intraoculaire.
Plusieurs essais cliniques ont déjà obtenu des résultats positifs en utilisant rAAV pour traiter Amaurose congénitale de Leber, montrant que le traitement est à la fois sûr et efficace Il n'y avait pas les événements indésirables graves, et les patients dans les trois études ont montré une amélioration de leur fonction visuelle telle que mesurée par un certain nombre de méthodes. Les méthodes utilisées variée parmi les trois essais, mais inclus les deux méthodes fonctionnels tels que l'acuité visuelle et la mobilité fonctionnelle ainsi que des mesures objectives qui sont moins sensibles à la polarisation, tels que la capacité de l'élève à répondre à la lumière et des améliorations à l'IRM fonctionnelle. Des améliorations ont été maintenus à long terme, avec des patients qui ont continué de bien faire après plus de 1,5 années.
L'architecture unique de la rétine et de son environnement relativement à l'abri privilégié aider ce processus. Jonctions serrées qui forment la barrière hémato-rétinienne séparer l'espace sous-rétinien de l'approvisionnement en sang, protégeant ainsi contre les microbes et les dégâts les plus à médiation immunitaire, et de renforcer sa capacité à répondre aux thérapies vectoriels médiation. L'anatomie très compartimenté de l'oeil facilite la livraison précise de suspensions de vecteurs thérapeutiques à des tissus spécifiques sous visualisation directe en utilisant des techniques de microchirurgie. Dans l'environnement protégé de la rétine, des vecteurs AAV sont capables de maintenir des niveaux élevés de l'expression du transgène dans l'épithélium pigmentaire rétinien (EPR), les photorécepteurs, les cellules ganglionnaires ou pour de longues périodes de temps après un seul traitement. En outre, l'œil et le système visuel peuvent être régulièrement et facilement surveillés pour la fonction visuelle et des changements structurels de la rétine après injections avec non invasive la technologie de pointe, tels que l'acuité visuelle, la sensibilité au contraste, fond auto-fluorescence (FAF), seuils visuels adaptés à l'obscurité, diamètres vasculaires, pupillométrie, électrorétinographie (ERG), ERG multifocale et tomographie par cohérence optique (OCT).
Cette stratégie est efficace contre un certain nombre de maladies de la rétine qui ont été étudiés, y compris les maladies néovasculaires qui sont caractéristiques de la dégénérescence maculaire liée à l'âge, la rétinopathie diabétique et la rétinopathie du prématuré. Étant donné que la régulation de la vascularisation de la rétine adulte implique un équilibre entre endogènes positifs facteurs de croissance, tels que le facteur de croissance endothelial vasculaire (VEGF) et des inhibiteurs de l'angiogenèse, tels que le facteur épithélium pigmentaire dérivé ( PEDF), l'expression de PEDF rAAV à médiation, l'angiostatine, et le récepteur de VEGF soluble sFlt-1, qui sont toutes les protéines anti-angiogéniques, se sont révélés réduire la formation de vaisseaux aberrant chez des modèles animaux. Étant donné que les thérapies géniques spécifiques ne peuvent pas facilement être utilisés pour traiter une fraction significative des patients atteints de dystrophie rétinienne, il est d'un intérêt majeur dans le développement d'une thérapie plus généralement applicables du facteur de survie. Les facteurs neurotrophiques ont la capacité de moduler la croissance neuronale au cours du développement pour maintenir des cellules existantes et pour permettre la récupération des populations neuronales blessé à l'oeil. AAV codant pour des facteurs neurotrophiques tels que le facteur de croissance des fibroblastes (FGF) et les membres de la famille GDNF photorécepteurs soit protégées de l'apoptose ou de ralentir la mort cellulaire.
Images supplémentaires

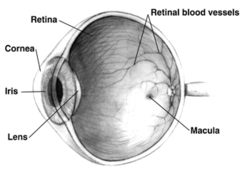
Les structures de l'œil marquées
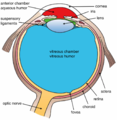
Une autre vue de l'œil et les structures de l'œil marqués