

Deinonychus
Acerca de este escuelas selección Wikipedia
SOS Children ha intentado que el contenido de Wikipedia más accesible por esta selección escuelas. Infantil SOS es la mayor donación de caridad del mundo niños huérfanos y abandonados de la oportunidad de la vida familiar.
| Deinonychus Rango temporal: Cretácico Temprano, 115-108Ma | |
|---|---|
 | |
| Esqueleto montado, Museo Field | |
| Clasificación científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Clase: | Reptilia |
| Orden: | Saurisquia |
| Familia: | † Dromaeosauridae |
| Clade: | † Eudromaeosauria |
| Subfamilia: | † Velociraptorinae |
| Género: | † Deinonychus Ostrom, 1969 |
| Especie: | † D. antirrhopus |
| Nombre binomial | |
| Antirrhopus Deinonychus Ostrom, 1969 | |
| Sinónimos | |
Velociraptor antirrhopus | |
Deinonychus (pron .: / d aɪ n ɒ n ɨ k ə s / dinámica- -ikəs NO; griego : δεινός, "terrible" y ὄνυξ, ὄνυχος genitivo 'garra') es un género de carnívoros dromeosáurido dinosaurios . Hay una especies descritas, Deinonychus antirrhopus. Estos dinosaurios, lo que podría crecer hasta 3,4 metros (11 pies) de largo, vivieron a principios del Cretácico Período, sobre 115-108 Hace millones de años (desde mediados Aptiano hasta principios Albian etapas). Los fósiles se han recuperado de la Estados de Montana, Wyoming, y Oklahoma, en rocas de la Cloverly Formación y Antlers Formación, aunque los dientes que pueden pertenecer a Deinonychus han encontrado mucho más al este en Maryland.
Paleontólogo El estudio de John Ostrom de Deinonychus a finales de 1960 revolucionó la manera como los científicos pensaban acerca de los dinosaurios, lo que lleva a la " renacimiento de los dinosaurios "y encender el debate sobre si los dinosaurios eran de sangre caliente o sangre fría. Antes de esto, la concepción popular de dinosaurios había sido uno de andar con paso pesado, gigantes reptiles. Ostrom observó el cuerpo pequeño, elegante, la postura horizontal, -rátidas como columna vertebral, y en especial las garras rapaces agrandadas en los pies, lo que sugiere un depredador ágil activo.
"Garra terrible" se refiere a la inusualmente grande garra, en forma de hoz en el segundo dedo de cada pata trasera. El fósil YPM 5205 conserva un grande, muy curvado ungueal. En la vida, arcosaurios tienen una vaina córnea sobre este hueso, que se extiende la longitud. Ostrom miró cocodrilos y aves garras y reconstruyó la garra para YPM 5205 como más de 120 milímetros (4,7 pulgadas) de largo. El nombre de la especie antirrhopus significa "contrapeso", que se refiere a la idea de Ostrom sobre la función de la cola. Al igual que en otros dromeosáuridos, la cola vértebras tienen una serie de tendones osificados y hueso muy alargada procesos. Estas características parecía hacer la cola en un contrapeso rígido, pero un fósil de la muy estrecha relación mongoliensis Velociraptor (IGM 100/986) tiene un esqueleto articulado cola que se curva lateralmente en una larga S-forma. Esto sugiere que, en la vida, la cola podría doblarse hacia los lados con un alto grado de flexibilidad.
Tanto en las formaciones Cloverly y Antlers, Deinonychus se han encontrado restos estrechamente asociada con las del ornithopod Tenontosaurus. Dientes descubierto asociados con especímenes Tenontosaurus implican fueron cazados, o al menos barridos sobre, por Deinonychus.
Descripción

Sobre la base de los especímenes más grandes conocidas, Deinonychus podría alcanzar al menos 3,4 metros (11 pies 2 pulgadas) de longitud, con una longitud cráneo de 410 mm (16,1 pulgadas), una altura de la cadera de 0,87 metros (2 pies 10 in), y una peso estimado de 73 kilogramos (160 libras). El cráneo estaba equipado con poderosas mandíbulas alineadas con alrededor de setenta, dientes como cuchillas curvas. Estudios del cráneo han progresado mucho durante las décadas. Ostrom reconstruyó los cráneos parciales, imperfectamente conservados que tuvo como triangular, ancha, y bastante similar a Allosaurus . El material adicional cráneo Deinonychus y especies estrechamente relacionadas que se encuentran con buena conservación tridimensional muestran que el paladar era más abovedado que se pensaba Ostrom, haciendo el hocico mucho más estrecho, mientras que el jugals estallaron en términos generales, dando mayor visión estereoscópica. El cráneo de Deinonychus era diferente de la de Velociraptor, sin embargo, en que tenía un techo del cráneo más robusto, como el de Dromaeosaurus, y no tenía las nasales deprimidas del Velociraptor. Tanto el cráneo y la mandíbula inferior tenían fenestras (aberturas del cráneo), que reduce el peso del cráneo. En Deinonychus, el fenestra antorbital, una abertura del cráneo entre el ojo y la fosa nasal, era particularmente grande.
Deinonychus poseía grandes "manos" ( manus) con tres garras en cada extremidad anterior. El primer dígito era más corta y la segunda era más larga. Cada pata trasera llevaba una garra en forma de hoz en el segundo dígito, lo que probablemente se utilizó durante la depredación.
No hay impresiones de piel nunca se han encontrado en asociación con fósiles de Deinonychus. Sin embargo, la evidencia sugiere que la Dromaeosauridae, incluyendo Deinonychus, tenía plumas. El género Microraptor es tanto filogenéticamente más antigua geológicamente y más primitivo que Deinonychus, y dentro de la misma familia. Múltiples fósiles de Microraptor preservan pennaceous, plumas de paletas como las de las aves modernas en los brazos, las piernas y la cola, junto con plumas secretas y curvas de nivel. Velociraptor es geológicamente más joven que Deinonychus, pero aún más estrechamente relacionados (dentro de la subfamilia Velociraptorinae, ver Clasificación , más abajo). Un ejemplar de Velociraptor se ha encontrado con las perillas de canilla en el cúbito. Perillas Quill son donde los ligamentos foliculares unidos, y son un indicador directo de plumas de aspecto moderno.
Clasificación

Antirrhopus Deinonychus es uno de los más conocidos dromeosáurido especies, y es un pariente cercano del menor Velociraptor , que se encuentra en más jóvenes, formaciones rocosas en edad Cretácico Superior en Asia Central. La clado forman se llama Velociraptorinae. El nombre subfamilia Velociraptorinae fue acuñado por primera vez por Rinchen Barsbold en 1983 y originalmente contenía el único género Velociraptor. Más tarde Phil Currie incluía la mayor parte de los dromeosáuridos. Dos géneros del Cretácico tardío, Tsaagan de Mongolia y la de América del Norte Saurornitholestes, también pueden ser familiares cercanos, pero este último es poco conocido y difícil de clasificar. Velociraptor y sus aliados están considerados como el uso de sus garras más que sus cráneos como herramientas de matar, en lugar de como dromaeosáuridos Dromaeosaurus, que tienen cráneos rechonchos. Junto con el troodontids, los dromaeosáuridos forman la Deinonychosauria clado, que es un taxón hermano de aves . Filogenéticamente, la Deinonychosauria representan el grupo de los dinosaurios no aviares más estrechamente relacionados con las aves.
Descubrimiento y denominación
Restos fosilizados de Deinonychus se han recuperado de la Cloverly Formación de Montana y Wyoming y en el más o menos contemporánea Formación de Antlers Oklahoma, en América del Norte . La formación Cloverly se ha fechado a finales del Aptiano hasta principios Etapas Albian de principios del Cretácico , alrededor de 115 a 108 Ma. Además, los dientes encontrados en el Arundel arcilla facies (mediados de Aptiano), de la Formación sobre el Potomac Atlantic Coastal Plain de Maryland puede ser asignado al género.
Los primeros restos fueron descubiertos en 1931 en el sur Montana, cerca de la ciudad de Billings. El líder del equipo, el paleontólogo Barnum Brown, se ocupa principalmente de excavación y preparación de los restos de la dinosaurio ornitópodo Tenontosaurus, pero en su informe de campo de la excavación a la Museo Americano de Historia Natural, informó el descubrimiento de un pequeño dinosaurio carnívoro cerca de un esqueleto Tenontosaurus ", pero encerrado en cal difícil de preparar." Él llama informalmente el animal "Daptosaurus agilis" e hizo los preparativos para describirla y tener el esqueleto, espécimen AMNH 3015, puesto en exhibición, pero nunca terminó este trabajo. Marrón trajo de la Formación Cloverly el esqueleto de un terópodo pequeño con dientes aparentemente de gran tamaño que informalmente llamado "Megadontosaurus". John Ostrom, la revisión de este material de décadas más tarde, se dio cuenta de que los dientes provenían de Deinonychus, pero el esqueleto de vino de un animal completamente diferente. Llamó a este esqueleto Microvenator.
Un poco más de treinta años después, en agosto de 1964, el paleontólogo John Ostrom dirigió una expedición desde el Museo Peabody de la Universidad de Yale que descubrió más material esquelético cerca Bridger. Expediciones durante los siguientes dos veranos al descubierto más de 1.000 huesos, entre los que había al menos tres personas. Desde la asociación entre los distintos huesos recuperados era débil, por lo que el número exacto de animales individuales representado imposible determinar correctamente, el espécimen tipo (YPM 5205) de Deinonychus se restringió al pie izquierdo completo y parcial del pie derecho que sin duda pertenecía a la misma persona. Las muestras restantes fueron catalogados en cincuenta entradas separadas en Yale Museo Peabody de Historia Natural.

Estudio posterior de Ostrom y Grant E. Meyer analiza su propio material, así como "Daptosaurus" de Brown en detalle y encontró que eran la misma especie. Ostrom publicó por primera vez sus conclusiones en febrero de 1969, dando todo el nuevo nombre se refería restos de Deinonychus antirrhopus. La nombre "antirrhopus" específica, desde ἀντίρροπος griego, significa "contrapeso" y se refiere a los efectos probables de una cola rígido. En julio de 1969, Ostrom publicó una muy extensa monografía sobre Deinonychus.
Aunque un gran número de huesos estaba disponible para 1969, muchas de las más importantes que faltaban o difícil de interpretar. Había pocos elementos postorbitales cráneo, no hay fémures, sin sacro, sin espoleta o esternón, vértebras que falta, y (Ostrom pensó) sólo un pequeño fragmento de un coracoides. Reconstrucción esquelética de Ostrom de Deinonychus incluido un hueso muy inusual pélvica - un pubis que era trapezoidal y plana, a diferencia de la de otros terópodos, pero que era la misma longitud que el isquion y que se encuentra justo al lado de él.
Otros resultados
En 1974, Ostrom publicó otra monografía sobre el hombro de Deinonychus en el que se dio cuenta de que el pubis que él había descrito era en realidad un coracoides - un elemento de hombro. En ese mismo año, otro ejemplar de Deinonychus, MCZ 4371, fue excavado en Montana por una expedición de la Universidad de Harvard, dirigido por Farish Jenkins. Este descubrimiento añade varios elementos nuevos: fémures bien conservados, pubes, un sacro, y mejores ilia, así como elementos de las pes y metatarso. Ostrom describió esta muestra y revisó su restauración esquelético de Deinonychus. Esta vez se mostró muy largo pubis, y Ostrom comenzó a sospechar que podrían haber sido incluso un poco retroversión como las de las aves.
Un esqueleto de Deinonychus, incluidos los huesos de la original (y más completa) AMNH 3015 muestra, se puede ver en la exhibición en el Museo Americano de Historia Natural, con otro ejemplar en exhibición en el Museo de Zoología Comparada de Universidad Harvard. Los especímenes del Museo Americano y Harvard son de una localidad diferente a los especímenes de Yale. Incluso estos dos montajes esqueléticos se carece de elementos, incluyendo el sterna, costillas esternales, espoleta, y gastralia.
Incluso después de todo el trabajo de Ostrom, varios pequeños bloques de material de lima-encajonado permanecieron sin preparación en depósito en el Museo de América. Estos consistieron sobre todo en huesos aislados y fragmentos de huesos, incluyendo la matriz original, o roca circundante en el que los especímenes fueron enterrados inicialmente. Un examen de estos bloques no preparadas por Gerald Grellet-Tinner y Peter Makovicky en 2000 reveló una interesante, característica por alto. Varios huesos largos, delgados identificadas en los bloques como tendones osificados (estructuras que ayudaron a endurecer la cola de Deinonychus) resultaron representar en realidad gastralia (costillas abdominales). Más significativamente, se descubrió un gran número de cáscaras fosilizadas previamente inadvertidas en la matriz de la roca que había rodeado a la muestra original Deinonychus.
En un informe posterior, más detallada sobre las cáscaras de huevo, Grellet-Tinner y Makovicky concluyeron que el huevo casi seguro que pertenecía a Deinonychus, lo que representa el primer huevo dromeosáurido ser identificado. Además, la superficie externa de una cáscara de huevo se encuentra en estrecho contacto con el gastralia lo que sugiere que podría tener Deinonychus meditado sus huevos. Esto implica que Deinonychus utiliza la transferencia de calor del cuerpo como un mecanismo para la incubación de los huevos, e indica una endotermia similar a las aves modernas. El estudio adicional por Gregory Erickson y sus colegas encuentra que este individuo tenía 13 o 14 años de edad en la muerte y su crecimiento se había estancado. A diferencia de otros terópodos en su estudio de los ejemplares encontrados en los huevos o nidos, que había terminado de crecer en el momento de su muerte.
Implicaciones
Descripción de Ostrom de Deinonychus en 1969 ha sido descrito como el descubrimiento más importante de la paleontología de dinosaurios en el siglo de mid-20th. El descubrimiento de este depredador ágil claramente activo hizo mucho para cambiar la concepción científica (y popular) de los dinosaurios y abrió la puerta a la especulación de que los dinosaurios podrían haber sido de sangre caliente. Este desarrollo se ha denominado la renacimiento de los dinosaurios. Varios años más tarde, Ostrom observó similitudes entre las patas delanteras de Deinonychus y el de las aves, una observación que le llevó a revivir la hipótesis de que las aves descienden de los dinosaurios. Cuarenta años después, esta idea es casi universalmente aceptada.
Debido a su anatomía extremadamente pájaro-como y estrecha relación con otros dromeosáuridos, los paleontólogos suponen que Deinonychus fue probablemente cubierto de plumas. Existe clara evidencia de fósiles de plumas modernas de estilo aviar durante varios dromaeosáuridos relacionados, incluyendo Velociraptor y Microraptor, aunque no hay evidencia directa es todavía conocido por sí Deinonychus. Cuando se realizan estudios de áreas tales como la amplitud de movimiento en las extremidades anteriores, los paleontólogos como Phil Senter han tomado la probable presencia de plumas de las alas (como presente en todos dromaeosaurios conocidos con impresiones de piel) en consideración.
Paleobiología y paleoecología
La evidencia geológica sugiere que Deinonychus habitado un llanuras inundables o hábitat swamplike. El paleoambiente tanto de la Formación Cloverly superior y el Antlers Formación, en el que queda de Deinonychus han sido encontrados, consistía en bosques tropicales o subtropicales, deltas y lagunas, no a diferencia de hoy Louisiana. Otros animales Deinonychus compartieron su mundo con incluir dinosaurios herbívoros como el blindado Sauropelta y los ornitópodos Zephyrosaurus y Tenontosaurus. En Oklahoma, el ecosistema de Deinonychus también incluyó el gran terópodo Acrocanthosaurus, el enorme saurópodo Sauroposeidon , la de cocodrilo Goniopholis, y el gar Lepisosteus. Si los dientes se encuentran en Maryland son los de Deinonychus, a continuación, sus vecinos se incluyen Astrodon y un nodosaur (llamado Priconodon) sólo se conoce de los dientes. La porción media de la Formación Cloverly rangos de edad de 115 ± 10 Ma cerca de la base a 108,5 ± 0,2 Ma cerca de la parte superior. En un estudio de 2001 realizado por Bruce Rothschild y otros paleontólogos, se examinaron 43 huesos de la mano y los 52 huesos del pie que se refiere el Deinonychus signos de fractura por estrés; no se encontró ninguno.
El comportamiento abusivo

Dientes Deinonychus encontrados en asociación con fósiles de la Tenontosaurus de dinosaurios ornitópodos son bastante comunes en la Formación Cloverly. Dos canteras se han descubierto fósiles que preservan bastante completos Deinonychus cerca fósiles Tenontosaurus. La primera, la cantera de Yale en los Cloverly de Montana, incluye numerosos dientes, cuatro adultos y un juvenil Deinonychus Deinonychus. La asociación de este número de Deinonychus esqueletos en una sola cantera sugiere que Deinonychus pudo haber alimentado de ese animal, y tal vez cazado. Ostrom y Maxwell incluso han utilizado esta información para especular que Deinonychus pudo haber vivido y cazado en manadas. El segundo ejemplo es la cantera de la Formación Antlers de Oklahoma. El sitio contiene seis esqueletos parciales de Tenontosaurus de varios tamaños, junto con un esqueleto parcial y muchos dientes de Deinonychus. Uno húmero tenontosaur incluso lleva lo que podría ser marcas de dientes de Deinonychus. Brinkman et al. (1998) señalan que Deinonychus tenía una masa de adultos de 70 a 100 kilogramos, mientras que tenontosaurs adultos eran 1.4 toneladas métricas. Un Deinonychus solitario no podía matar a un tenontosaur adulto, lo que sugiere que la caza paquete es posible.
Un estudio realizado en 2007 por Roach y Brinkman ha puesto en duda el paquete de comportamiento de caza cooperativa de Deinonychus, en base a lo que se conoce de la caza carnívoro moderna y la tafonomía de sitios tenontosaur. Moderno arcosaurios ( aves y cocodrilos ) y dragones de Komodo muestran poca caza cooperativa; en cambio, por lo general son bien cazadores solitarios, o se sienten atraídos por las canales previamente asesinados, donde se produce mucho conflicto entre individuos de la misma especie. Por ejemplo, en situaciones en las que grupos de dragones de Komodo están comiendo juntos, los individuos más grandes comen primero y atacarán Komodos más pequeños que tratan de comer; si el animal más pequeño es asesinado, es canibalizado. Cuando se aplica esta información a los sitios tenontosaur, parece que lo que se encuentra es consistente con Deinonychus tener una estrategia de alimentación komodo- o cocodrilo-como. Deinonychus restos óseos encontrados en estos sitios son de subadultos, con partes faltantes consistente con haber sido comido por otra Deinonychus. Por otra parte, un artículo de Li et al., Describe los sitios de pista con una separación pie similar y rastros paralelos, lo que implica el comportamiento de embalaje gregaria en lugar de comportamiento de alimentación no coordinada.
En 2011, Denver Fowler y sus colegas sugirieron un nuevo método por el cual Deinonychus y otros dromaeosaurios pueden haber capturado y presas restringido. Este modelo, conocido como la "presa rapaz moderación" (RPR) modelo de depredación, propone que Deinonychus mató a su presa de una manera muy similar a la existente accipitrid aves rapaces : al saltar sobre su presa, clavándolo en virtud de su peso corporal, y agarrando firmemente con las grandes garras en forma de hoz. Como accipitrids, la dromaeosaur comenzaría entonces a alimentarse de los animales en vida, hasta que finalmente murió a causa de la pérdida de sangre e insuficiencia orgánica. Esta propuesta se basa principalmente en las comparaciones entre la morfología y las proporciones de los pies y las piernas de los dinosaurios, a varios grupos de aves rapaces existentes con comportamientos depredadores conocidos. Fowler encontró que los pies y las piernas de los dinosaurios, más semejantes a los de las águilas y halcones , especialmente en términos de tener una segunda garra ampliada y una gama similar de captar el movimiento. Sin embargo, el corto metatarso y la fuerza del pie habrían sido más similar a la de los búhos . El método RPR de depredación sería coherente con otros aspectos de la anatomía Deinonychus 's, como la mandíbula y el brazo morfología inusual. Las armas fueron probablemente cubiertos de plumas largas, y pueden haber sido utilizados como aleteo estabilizadores para el equilibrio mientras lo alto de un animal de presa luchando, junto con la cola tiesa contrapeso. Las mandíbulas, que se cree que han tenido una fuerza de mordida comparativamente débil, habría sido útil para comer presas vivas, pero no tan útil para la rápida expedición, por la fuerza de la presa (ya que los animales modernos que hacen esto tienen especializaciones únicas que dromaeosaurios carecían). Estas adaptaciones predadoras que trabajan juntos también pueden tener implicaciones para el origen de aleteo en paravians.
Fuerza de mordida

Estimaciones de la fuerza de mordida de Deinonychus se produjeron por primera vez en 2005, basado en la musculatura de la mandíbula reconstruida. Este estudio concluyó que Deinonychus probablemente tenía una fuerza máxima de mordedura sólo el 15% de la de la moderna Cocodrilo americano. Un estudio de 2010 por Paul y sus colegas Gignac intentó estimar la fuerza de mordida basada directamente en marcas de pinchazos Deinonychus diente recién descubiertos en los huesos de un Tenontosaurus. Estas marcas de pinchazos provenían de una gran persona, y siempre la primera evidencia de que las grandes Deinonychus podrían picar a través del hueso. Usando las marcas de dientes, el equipo de Gignac fueron capaces de determinar que la fuerza de mordedura de Deinonychus fue significativamente mayor que estudios anteriores habían estimado por estudios biomecánicos solo. Ellos encontraron la fuerza de mordida de Deinonychus ser entre 4100 y 8200 newtons, mayor que viven mamíferos carnívoros como la hiena , y equivalente a un cocodrilo de tamaño similar.
Gignac y sus colegas también observaron, sin embargo, que las marcas de punción de médula de Deinonychus son relativamente raras, ya diferencia de los terópodos más grandes con muchas marcas de pinchazos conocidos como Tyrannosaurus , Deinonychus probablemente no muerden frecuentemente a través de o comen los huesos. En cambio, es probable que utilizaron su alta fuerza de mordida en la defensa o la captura de presas, en lugar de alimentar.
La función del miembro

A pesar de ser la característica más distintiva de Deinonychus, la forma y la curvatura de la garra falciformes varía entre los especímenes. El espécimen tipo descritos por Ostrom en 1969 tiene una garra falciforme fuertemente curvada, mientras que una muestra más nuevos descritos en 1976 tenían una garra con curvatura mucho más débil, más similares en perfil con las garras "normales" en los dedos restantes. Ostrom sugirió que esta diferencia en el tamaño y forma de las garras de hoz podría ser debido a la variación individual, sexual, o relacionada con la edad, pero admitió que no podía estar seguro.
Hay evidencia anatómica y rastrillada que este talon se levantó del suelo, mientras que el dinosaurio caminaba sobre el tercer y cuarto dedo del pie.
Ostrom sugiere que Deinonychus podía patear con la garra de la hoz para cortar y tajar a su presa. Algunos investigadores sugieren que la garra se utilizó para destripar grande dinosaurios ceratopsian. Otros estudios han sugerido que las garras de hoz no estaban acostumbrados a recortar sino para ofrecer pequeñas puñaladas a la víctima. Manning et al. (2005) realizó pruebas sobre una réplica robótica que justamente coincidía con la anatomía de Deinonychus y el Velociraptor , y utiliza cilindros hidráulicos para hacer que el robot lograr un canal de cerdo. En estas pruebas, las garras hechas sólo pinchazos superficiales y no podían cortar o recortar. Los autores sugirieron que las garras habrían sido más eficaces en la escalada que en asestar golpes matando.
Ostrom comparó Deinonychus al avestruz y emús . Señaló que las especies de aves pueden causar una lesión grave con la gran garra en el segundo dedo del pie. El casuario tiene garras de hasta 125 milímetros (4,9 pulgadas) de largo. Ostrom citó Gilliard (1958) al decir que pueden romper un brazo o destripar a un hombre. Kofron (1999 y 2003) estudió 241 ataques casuario documentados y encontró que uno dos perros humana y habían muerto, pero no hay evidencia de que los casuarios pueden destripar o desmembrar otros animales. Casuarios utilizan sus garras para defenderse, para atacar a los animales amenazantes, y en despliegues agonísticos, como la amenaza de visualización de Arco. La chuña también tiene una segunda garra pies ampliada, y lo utiliza para desgarrar pequeñas presas para tragar. En 2011, un estudio sugiere que la garra falciforme probablemente habría sido utilizada para precisar presa mientras que morder, y no como un arma cortante.
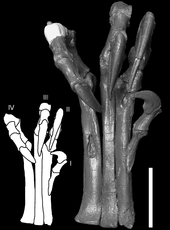
Estudios biomecánicos por Ken Carpenter, en 2002 confirmó que la función más probable de las patas delanteras en la depredación estaba agarrando, ya que sus grandes esfuerzos habrían permitido alcanzar ya que para la mayoría de los terópodos. El bastante grande y alargada coracoides, indicando poderosos músculos en las extremidades anteriores, fortaleció aún más esta interpretación. Estudios biomecánicos de Carpenter utilizando hueso yesos también mostraron que Deinonychus no podía doblar sus brazos contra su cuerpo como un pájaro ("plegado aviar"), en contra de lo que se infiere de las 1.985 descripciones anteriores por Jacques Gauthier y Gregory S. Paul en 1988.
Los estudios realizados por Phil Senter en 2006 indicó que las extremidades anteriores Deinonychus podrían utilizarse no sólo para agarrar, sino también para agarrando objetos hacia el pecho. Si Deinonychus tenía dedos de plumas y alas, las plumas se habrían limitado el rango de movimiento de las extremidades anteriores hasta cierto punto. Por ejemplo, cuando Deinonychus extendió su brazo hacia adelante, la "palma 'de la mano gira automáticamente a una posición orientada hacia arriba. Esto habría causado un ala para bloquear la otra si ambas extremidades anteriores se extendieron al mismo tiempo, lo que lleva a concluir que Senter agarrando objetos en el pecho serían sólo se han logrado con un brazo a la vez. La función de los dedos también se habría limitado por las plumas; por ejemplo, sólo el tercer dígito de la mano podría haber sido empleado en actividades tales como grietas de sondeo para las pequeñas presas, y sólo en una posición perpendicular al ala principal. Alan Gishlick, en un estudio de 2001 de la mecánica de las extremidades anteriores Deinonychus, encontró que incluso si las grandes plumas de las alas estaban presentes, la capacidad de agarre de la mano no se habría visto obstaculizada de manera significativa; más bien, agarre se habría logrado perpendicular al ala, y objetos probablemente habría estado en manos de las dos manos al mismo tiempo de una manera "abrazo de oso", hallazgos que han sido apoyados por los estudios posteriores de las extremidades anteriores Carpenter y Senter.
Parsons y Parsons han demostrado que los juveniles y subadultos ejemplares de Deinonychus muestran algunas diferencias morfológicas con los adultos. Por ejemplo, los brazos de los ejemplares más jóvenes fueron proporcionalmente más que los de los adultos, una posible indicación de la diferencia de comportamiento entre jóvenes y adultos. Otro ejemplo de esto podría ser la función de las garras de pedal. Parsons y Parsons han sugerido que la curvatura de la garra (que Ostrom [1976] ya había mostrado fue diferente entre las muestras) quizás fue mayor para Deinonychus juvenil, ya que esto podría ayudar a trepar en los árboles, y que las garras se hizo más recto que el animal se volvió más viejo y comenzó a vivir únicamente en el suelo. Esto se basa en la hipótesis de que algunos pequeños dromaeosáuridos utilizan sus garras pedal para subir.
Velocidad

Dromaeosaurids, especialmente Deinonychus, se representan a menudo como animales inusualmente rápidos de marcha en los medios populares, y el propio Ostrom especularon que Deinonychus era veloz en su descripción original. Sin embargo, cuando se describió por primera vez, no se había encontrado una pierna completa de Deinonychus, y la especulación de Ostrom sobre la longitud de la fémur (hueso superior de la pierna) más adelante se demuestre que ha sido una sobreestimación. En un estudio posterior, Ostrom observó que la relación del fémur a la tibia (hueso de la pierna inferior) no es tan importante en la determinación de la velocidad como la longitud relativa del pie y la pierna inferior. En las aves veloces modernos, como el avestruz , la relación pies tibia es 0,95. En dinosaurios inusualmente rápido de marcha, como Struthiomimus , la proporción es de 0,68, pero en Deinonychus la relación es de 0,48. Ostrom declaró que "la única conclusión razonable" es que Deinonychus no fue particularmente rápido en comparación con otros dinosaurios, y ciertamente no tan rápido como aves no voladoras modernas.
La baja del pie a la proporción inferior de la pierna en Deinonychus se debe en parte a un inusualmente corto metatarso (huesos del pie superior). La relación es en realidad mayor en los individuos más pequeños que en las grandes. Ostrom sugirió que el corto metatarso puede estar relacionado con la función de la garra de hoz, y se utiliza el hecho de que parece que se corta como individuos de edades comprendidas como soporte para esto. Interpretó todas estas características - el corto segundo dedo con garra agrandada, corto metatarso, etc. - como el apoyo a la utilización de la pata trasera como arma ofensiva, donde la garra falciforme golpearía hacia abajo y hacia atrás, y la pierna hacia atrás y hacia abajo al mismo tiempo, la roza y lagrimeo en la presa. Ostrom sugirió que el corto metatarso reduce el estrés general en los huesos de las piernas durante un ataque de este tipo, y se interpreta la disposición inusual de las inserciones musculares en la pierna Deinonychus como apoyo a su idea de que se utilizó un conjunto diferente de los músculos en la carrera depredadora que en pie o correr. Por lo tanto, Ostrom concluyó que las patas de Deinonychus representaba un equilibrio entre las adaptaciones de funcionamiento necesarios para un depredador ágil, y las características de reducción de estrés para compensar su arma única pie.
En su estudio de huellas de dinosaurios de Canadá 1981, Richard Kool produjo estimaciones de la velocidad para caminar en bruto basado en varios rastros realizados por diferentes especies en el Gething Formación de Columbia Británica. Kool estima uno de estos rastros, lo que representa el icnoespecie Irenichnites gracilis (que puede haber sido hecha por Deinonychus), para tener una velocidad de caminata de 10,1 kilómetros por hora (6 millas por hora).
Huevos

La identificación, en 2000, de un huevo de Deinonychus probable asociado con uno de los ejemplares originales permiten la comparación con otros dinosaurios terópodos en términos de estructura de huevo, anidación y reproducción. En su examen de la muestra de 2006, Grellet-Tinner y Makovicky examinaron la posibilidad de que el dromeosáurido había estado alimentando en el huevo, o que los fragmentos de huevos habían sido asociados con el esqueleto Deinonychus por casualidad. Ellos rechazaron la idea de que el huevo había sido una comida para el terópodo, señalando que los fragmentos se intercalan entre las costillas del vientre y los huesos de la extremidad anterior, lo que hace imposible que representaban contenido del estómago del animal. Además, la manera en que el huevo había sido aplastada y fragmentado indicó que había sido intacta en el momento del entierro, y fue roto por el proceso de fosilización. La idea de que el huevo se asocia aleatoriamente con el dinosaurio también se encontró que era poco probable; los huesos que rodean el huevo no habían sido dispersada o desarticulada, pero se mantuvo relativa bastante intacta a sus posiciones en la vida, lo que indica que el área alrededor y que incluye el óvulo no fue perturbada durante la conservación. El hecho de que estos huesos fueron costillas vientre ( gastralia), que es muy raro encontrar articulado, con el apoyo de esta interpretación. Toda la evidencia, según Grellet-Tinner y Makovicky, indican que el huevo estaba intacto debajo del cuerpo del Deinonychus cuando fue enterrado. Es posible que este representa el comportamiento de crianza o de anidación en Deinonychus similar a la observada en el relacionado troodontids y oviraptóridos, o que el huevo era, de hecho, dentro de la oviducto cuando el animal murió.
El examen de la microestructura del huevo Deinonychus confirma que perteneció a un terópodo, ya que comparte características con otros huevos terópodos conocidos y muestra diferencias con ornitisquio y huevos de saurópodos. En comparación con otra terópodos maniraptoran, el huevo de Deinonychus es más similar a las de oviraptóridos que a los de troodontids, a pesar de los estudios que muestran este último están más estrechamente relacionados con dromaeosáuridos como Deinonychus. Mientras que el huevo fue demasiado mal aplastado para determinar con precisión su tamaño, Grellet-Tinner y Makovicky estima un diámetro de aproximadamente 7 cm (2,7 pulgadas) sobre la base de la anchura de la canal pélvico a través del cual el huevo tuvo que han pasado. Este tamaño es similar al diámetro 7,2 cm de la más grande Citipati (un oviraptorid) huevos; Citipati y Deinonychus también compartió el mismo tamaño del cuerpo en general, el apoyo a esta estimación. Además, los espesores de Citipati y Deinonychus cáscaras de huevo son casi idénticos, y desde grosor de la cáscara se correlaciona con el volumen de huevos, este también apoya la idea de que los huevos de estos dos animales eran aproximadamente del mismo tamaño.
Patología
La segunda falange del segundo dedo del pie en el YPM espécimen 5205 tiene una fractura curada.