

Mitocondria
Sabías ...
Esta selección wikipedia ha sido elegido por los voluntarios que ayudan Infantil SOS de Wikipedia para esta Selección Wikipedia para las escuelas. SOS Children ha cuidado de niños en África durante cuarenta años. ¿Puedes ayudar a su trabajo en África ?

En biología celular, una mitocondria (mitocondrias plural) es una membrana encerrada- orgánulo encuentra en la mayoría de los eucariotas células . Estos orgánulos varían de 1-10 micrómetros ( m) en tamaño. Las mitocondrias se describen a veces como "plantas de energía celulares", ya que generan la mayor parte del suministro de la célula de trifosfato de adenosina (ATP), que se utiliza como una fuente de energía química. Además del suministro de energía celular, las mitocondrias están involucrados en una variedad de otros procesos, tales como de señalización, la diferenciación celular, la muerte celular, así como el control de la ciclo celular y el crecimiento celular. Las mitocondrias se han implicado en varias enfermedades humanas, incluyendo trastornos mentales, disfunción cardíaca, y puede desempeñar un papel en el proceso de envejecimiento. La palabra proviene de la mitocondria griega μίτος o mitos, hilo + χονδρίον o khondrion, gránulo. Su ascendencia no se entiende completamente, pero, de acuerdo con la teoría endosimbiótica, las mitocondrias son descendientes de la antigua bacterias, que fueron devorados por los ancestros de las células eucariotas hace más de mil millones de años.
Varias características hacen que las mitocondrias único. El número de mitocondrias en una célula varía ampliamente según el organismo y tipo de tejido. Muchas células tienen sólo una única mitocondria, mientras que otros pueden contener varios miles de mitocondrias. El orgánulo se compone de compartimentos que llevan a cabo funciones especializadas. Estos compartimentos o regiones incluyen la membrana externa, la espacio intermembrana, la membrana interna, y el crestas y matriz. Las proteínas mitocondriales varían dependiendo de los tejidos y especies. En humanos, se identificaron 615 tipos diferentes de proteínas de la mitocondria cardiaca; mientras que en Se informó, 940 proteínas codificadas por los genes murinos distintos. Proteoma mitocondrial se cree que está regulada dinámicamente. Aunque la mayor parte del ADN de una célula está contenido en el núcleo de la célula, la mitocondria tiene su propia independiente genoma. Además, su ADN muestra similitud sustancial a bacteriana genomas.
Estructura

A mitocondria contiene membranas internas y externas compuestas de bicapas de fosfolípidos y proteínas . Las dos membranas, sin embargo, tienen propiedades diferentes. Debido a esta organización de doble membraned, hay cinco compartimentos diferenciados dentro de la mitocondria. No es el la membrana mitocondrial externa, la espacio intermembrana (el espacio entre las membranas externa e interna), el membrana mitocondrial interna, la espacio crestas (formado por invaginaciones de la membrana interna), y el matriz (espacio dentro de la membrana interna).
Membrana externa
La membrana mitocondrial externa, que encierra la totalidad orgánulo, tiene una proteína-a- relación de fosfolípido similar a la de la membrana plasmática eucariota (alrededor de 1: 1 en peso). Contiene un gran número de proteínas integrales llamados porinas. Estos porinas forman canales que permiten que las moléculas de 5000 Daltons o menos en su peso molecular a libremente difundir desde un lado de la membrana al otro. Las proteínas más grandes también pueden entrar en la mitocondria si una secuencia de señalización en su N-terminal se une a un gran proteína multisubunit llama translocasa de la membrana externa, que luego los mueve activamente a través de la membrana. La interrupción de la membrana externa permite proteínas en el espacio intermembrana a filtrarse en el citosol, lo que conduce a una muerte segura celular.
Espacio intermembrana
La espacio intermembrana es el espacio entre la membrana exterior y la membrana interior. Debido a que la membrana externa es libremente permeable a pequeñas moléculas, las concentraciones de moléculas pequeñas tales como iones y azúcares en el espacio intermembrana es la misma que la citosol. Sin embargo, como proteínas grandes deben tener una secuencia de señalización específico para ser transportados a través de la membrana externa, la composición de proteínas de este espacio es diferente de la composición de proteínas de la citosol. Una proteína que se localiza en el espacio intermembrana de esta manera es citocromo c.
Membrana interna
La membrana mitocondrial interna contiene proteínas con cuatro tipos de funciones:
- Aquellos que realizan la reacciones redox de la fosforilación oxidativa
- ATP sintasa, que genera ATP en la matriz
- Proteínas de transporte específicas que regulan paso metabolito dentro y fuera de la matriz
- Maquinaria de importación de proteínas.
Contiene más de 100 diferentes polipéptidos, y tiene una relación muy alta de proteína a fosfolípido (más de 3: 1 en peso, que es aproximadamente 1 proteína para 15 fosfolípidos). La membrana interna es el hogar de alrededor de un quinto del total de proteínas en una mitocondria. Además, la membrana interna es rica en un fosfolípido inusual, cardiolipina. Este fosfolípido fue descubierto originalmente en corazones de carne en 1942, y suele ser característico de las membranas plasmáticas mitocondriales y bacterianos. Cardiolipina contiene cuatro ácidos grasos en lugar de dos y puede ayudar a hacer que la membrana interna impermeable. A diferencia de la membrana exterior, la membrana interior no contiene porinas y es altamente impermeable a todas las moléculas. Casi todos los iones y moléculas requieren transportadores de membrana especiales para entrar o salir de la matriz. Las proteínas se transportan en la matriz a través de la translocasa del complejo de la membrana interior (TIM) o por medio de Oxa1. Además, hay un potencial de membrana a través de la membrana interna formada por la acción de las enzimas de la cadena de transporte de electrones.
Crestas

La membrana mitocondrial interna está compartimentada en numerosos crestas, que amplían la superficie de la membrana mitocondrial interna, mejorando su capacidad de producir ATP. Estos son pliegues azar no simples sino más bien invaginaciones de la membrana interna, que pueden afectar global función quimiosmótica. En típico mitocondrias de hígado, por ejemplo, el área de superficie, incluyendo crestas, es aproximadamente cinco veces mayor que la de la membrana externa. Las mitocondrias de células que tienen una mayor demanda de ATP, tales como las células musculares, contiene más crestas de las mitocondrias de hígado típicos.
Matriz
La matriz es el espacio encerrado por la membrana interna. Contiene aproximadamente 2/3 de la proteína total en una mitocondria. La matriz es importante en la producción de ATP con la ayuda de la sintasa de ATP contenida en la membrana interna. La matriz contiene una mezcla altamente concentrada de cientos de enzimas, mitocondrial especial ribosomas, tRNA, y varias copias de la ADN mitocondrial genoma. De las enzimas, las funciones principales incluyen la oxidación de piruvato y ácidos grasos , y el ciclo del ácido cítrico.
Las mitocondrias tienen su propio material genético, y la maquinaria para fabricar su propio ARN y proteínas (véase: la biosíntesis de proteínas). Una secuencia de ADN mitocondrial humano publicado reveló 16569 pares de bases que codifican 37 genes totales, 24 ARNt y genes de ARNr y 13 genes de péptidos. El 13 mitocondrial péptidos en los seres humanos están integrados en la membrana mitocondrial interna, junto con las proteínas codificadas por genes que residen en el huésped celular de núcleo.
Organización y distribución
Las mitocondrias se encuentran en casi todos los eucariotas . Ellos varían en número y ubicación según el tipo de célula. Un número considerable de las mitocondrias son en el hígado, con alrededor de 1000-2000 mitocondrias por célula que componen 1 / quinto del volumen celular. Las mitocondrias se encuentran ubicado entre miofibrillas de muscular o envuelto alrededor de la esperma flagelo. A menudo forman una compleja red de ramificación 3D dentro de la célula con el citoesqueleto. La asociación con el citoesqueleto determina la forma mitocondrial, lo que puede afectar a la función también. La evidencia reciente sugiere vimentina, uno de los componentes del citoesqueleto, es crítico para la asociación con el citoesqueleto.
Función
Los papeles más destacados de la mitocondria son su producción de ATP y la regulación de celulares metabolismo. El conjunto central de reacciones implicadas en la producción de ATP se conocen colectivamente como la ciclo del ácido cítrico. Sin embargo, la mitocondria tiene muchas otras funciones además de la producción de ATP.
Conversión de energía
Un papel dominante para la mitocondria es la producción de ATP , como se refleja en el gran número de proteínas en la membrana interna para esta tarea. Esto se realiza mediante la oxidación de los principales productos de la glucosa , piruvato, y NADH, que se producen en el citosol. Este proceso de la respiración celular, también conocido como la respiración aeróbica, es dependiente de la presencia de oxígeno . Cuando el oxígeno es limitado, los productos glucolíticas se metabolizan por la respiración anaeróbica, un proceso que es independiente de las mitocondrias. La producción de ATP a partir de glucosa tiene un aproximadamente 13 veces mayor rendimiento durante la respiración aeróbica en comparación con la respiración anaerobia.
Piruvato: el ciclo del ácido cítrico
Cada molécula de piruvato producido por glucólisis es transportada activamente a través de la membrana mitocondrial interna, y en la matriz donde se oxidada y combinado con coenzima A para formar CO 2, acetil-CoA, y NADH.
La acetil-CoA es el sustrato principal para entrar en el ciclo del ácido cítrico, también conocido como el ciclo del ácido tricarboxílico (TCA) o ciclo de Krebs. Las enzimas del ciclo del ácido cítrico se encuentran en la matriz mitocondrial, con la excepción de succinato deshidrogenasa, que está unido a la membrana mitocondrial interna como parte del complejo II. El ciclo del ácido cítrico oxida el acetil-CoA a dióxido de carbono, y, en el proceso, produce cofactores reduce (tres moléculas de NADH y una molécula de FADH 2) que son una fuente de electrones para el cadena de transporte de electrones, y una molécula de GTP (que se convierte fácilmente en un ATP).
NADH y FADH 2: la cadena de transporte de electrones
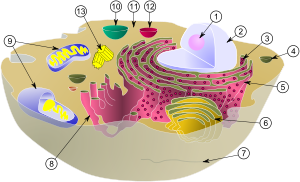
(1) nucléolo
(2) núcleo
(3) los ribosomas (pequeños puntos)
(4) vesícula
(5) en bruto retículo endoplásmico (ER)
(6) Aparato de Golgi
(7) Citoesqueleto
(8) RE liso
(9) las mitocondrias
(10) vacuola
(11) citoplasma
(12) lisosoma
(13) centríolos dentro centrosoma
La energía redox de NADH y FADH 2 se transfiere al oxígeno (O 2) en varios pasos a través de la cadena de transporte de electrones. Estas moléculas ricas en energía se producen dentro de la matriz a través del ciclo del ácido cítrico, pero también se producen en el citoplasma por glucólisis. La reducción de los equivalentes desde el citoplasma se puede importar a través de la sistema de lanzadera de malato-aspartato proteínas antiporter o se alimentan en la cadena de transporte de electrones mediante un lanzadera glicerol fosfato. Complejos de proteínas en la membrana interna ( NADH deshidrogenasa, citocromo c reductasa, y citocromo oxidasa c) realizar la transferencia y la liberación gradual de la energía se utiliza para bombear protones (H +) en el espacio intermembrana. Este proceso es eficiente, pero un pequeño porcentaje de electrones puede prematuramente reducir el oxígeno, formando especies de oxígeno reactivas tales como superóxido. Esto puede causar estrés oxidativo en la mitocondria y puede contribuir a la disminución de la función mitocondrial asociada con el proceso de envejecimiento.
Como la concentración de protones aumentos en el espacio intermembrana, un fuerte gradiente electroquímico se establece a través de la membrana interna. Los protones pueden volver a la matriz a través de la Complejo ATP sintasa, y su potencial de energía se utiliza para sintetizar ATP a partir de ADP y fosfato inorgánico (P i). Este proceso se llama quimiosmosis, y fue descrito por primera vez por Peter Mitchell que fue galardonado con el 1978 Premio Nobel de Química por su trabajo. Más tarde, parte del Premio Nobel 1997 de Química fue otorgado a Paul D. Boyer y John E. Walker para su esclarecimiento del mecanismo de trabajo de la ATP sintasa.
La producción de calor
Bajo ciertas condiciones, los protones pueden volver a entrar en la matriz mitocondrial sin contribuir a la síntesis de ATP. Este proceso se conoce como fuga de protones o desacoplamiento mitocondrial y es debido a la La difusión facilitada de protones en la matriz. El proceso resulta en la energía potencial descontrolado del gradiente electroquímico de protones que se liberan en forma de calor. El proceso está mediado por un canal de protones llamada thermogenin, o UCP1. Thermogenin es un 33k Proteína Da descubrió por primera vez en 1973. thermogenin se encuentra principalmente en tejido adiposo marrón o grasa marrón, y es responsable de la termogénesis sin temblor. El tejido adiposo marrón se encuentra en los mamíferos, y se encuentra en sus niveles más altos en la vida temprana y en los animales que hibernan. En los seres humanos, el tejido adiposo marrón está presente al nacer y disminuye con la edad.
Almacenamiento de iones de calcio
Las concentraciones de calcio libre en la célula pueden regular una serie de reacciones y es importante para transducción de señales en la célula. Las mitocondrias puede transitoriamente tienda de calcio, un proceso que contribuye a la homeostasis de la célula de calcio. De hecho, su capacidad para tomar rápidamente en calcio para su posterior liberación hace que sean muy buenos "buffers" citosólicos de calcio. La retículo endoplásmico (ER) es el sitio de almacenamiento más importante de calcio, y hay una interacción significativa entre la mitocondria y ER con respecto al calcio. El calcio se absorbe en el por una matriz de calcio uniporter en el membrana mitocondrial interna. Es impulsado principalmente por el mitocondrial potencial de membrana. La liberación de este calcio de nuevo en el interior de la célula puede ocurrir a través de una proteína de intercambio de sodio-calcio o por vías "calcio de liberación inducida por calcio". Esto puede iniciar picos de calcio o las ondas de calcio con grandes cambios en el potencial de membrana. Estos pueden activar una serie de proteínas segundo sistema de mensajería que pueden coordinar procesos como la liberación de neurotransmisores en las células nerviosas y la liberación de las hormonas en las células endocrinas.
Funciones adicionales
Las mitocondrias juegan un papel central en muchos otros tareas metabólicas, tales como:
- Reglamento de la potencial de membrana
- La apoptosis muerte celular programada
- Excitotoxicidad mediada por glutamato lesión neuronal
- Regulación de la proliferación celular
- Reglamento de celular metabolismo
- Cierto reacciones de síntesis del grupo hemo (véase también: porfirina)
- La síntesis de esteroides.
Algunas funciones mitocondriales se realizan sólo en tipos específicos de células. Por ejemplo, las mitocondrias en las células del hígado contienen enzimas que les permiten desintoxicar el amoníaco , un producto de desecho del metabolismo de proteínas. Una mutación en los genes que regulan cualquiera de estas funciones puede resultar en enfermedades mitocondriales.
Origen
Las mitocondrias tienen muchas características en común con procariotas. Como resultado, se cree que se deriva originalmente de procariotas endosymbiotic.
Una mitocondria contiene ADN, que se organiza en varias copias de un solo cromosoma circular. Este cromosoma mitocondrial contiene genes para ribosomas, y los veintiún de ARNt necesario para la traducción de ARN mensajero en proteína. La estructura circular también se encuentra en procariotas, y la similitud se extiende por el hecho de que el ADN mitocondrial se organiza con una variante código genético similar a la de Proteobacteria. Esto sugiere que su antepasado, el llamado proto-mitocondria, fue miembro de la Proteobacteria. En particular, el proto-mitocondria fue probablemente relacionado con el rickettsia. Sin embargo, la relación exacta del ancestro de la mitocondria a la alfa-proteobacterias y si las mitocondrias se formó al mismo tiempo o después de que el núcleo, sigue siendo controvertido.
Los ribosomas codificadas por el ADN mitocondrial son similares a los de las bacterias en tamaño y estructura. Ellos se parecen mucho a la bacteriana Ribosoma 70S y no el 80S ribosomas citoplásmicos que están codificados por ADN nuclear.
La relación endosimbiótica de las mitocondrias con sus células huésped fue popularizado por Lynn Margulis. La hipótesis endosimbiótica sugiere que las mitocondrias descendiente de bacterias que de algún modo sobrevivieron endocitosis por otra célula, y llegó a ser incorporado en el citoplasma. La capacidad de estas bacterias para llevar a cabo la respiración en las células huésped que había confiado en glicolisis y fermentación habría proporcionado una considerable ventaja evolutiva. De manera similar, las células huésped con bacterias simbióticas capaces de fotosíntesis también habrían tenido una ventaja. La incorporación de simbiontes habría aumentado el número de entornos en los que las células podrían sobrevivir. Esta relación simbiótica probablemente desarrolló hace 1,7-2000000000 años.
Unos grupos de eucariotas unicelulares carecen de mitocondrias: la microsporidians, metamonads, y Archamoebae. Estos grupos aparecen como los eucariotas más primitivos en árboles filogenéticos construidos usando información rRNA, lo que sugiere que se presentaron ante el origen de las mitocondrias. Sin embargo, esto es ahora conocido por ser un artefacto de larga rama atracción - que se derivan los grupos y retienen genes u orgánulos derivados de las mitocondrias (por ejemplo, mitosomes y hidrogenosomas).
Genoma
El genoma mitocondrial humano es una circular de ADN molécula de alrededor de 16 kilobases. Codifica 37 genes: 13 para subunidades de complejos respiratorios I, III, IV, y V, 22 para mitocondrial tRNA, y 2 para rRNA. Una mitocondria puede contener nueve y cincuenta y ocho copias de su ADN.
Al igual que en los procariotas, hay una muy alta proporción de ADN que codifica y una ausencia de repeticiones. Genes mitocondriales son transcrito como transcripciones multigenic, que se escinden y poliadenilado para producir madura mRNAs. No todas las proteínas necesarias para la función mitocondrial están codificadas por el genoma mitocondrial; la mayoría están codificadas por genes en el núcleo de la célula y las proteínas correspondientes importados en la mitocondria. El número exacto de genes codificados por el núcleo y el genoma mitocondrial difiere entre especies. En general, los genomas mitocondriales son circulares, aunque se ha informado excepciones. También, en general, el ADN mitocondrial carece intrones, como es el caso en el genoma mitocondrial humano; Sin embargo, los intrones se han observado en algunos de ADN mitocondrial eucariota, tal como el de la levadura y protistas, incluyendo Dictyostelium discoideum.
Mientras que pequeñas variaciones en el código estándar habían predicho anteriormente, ninguno fue descubierto hasta 1979, cuando los investigadores estudiar genes mitocondriales humanos determinaron que utilizaron un código alternativo. Muchos ligeras variantes se han descubierto desde entonces, incluyendo diversos códigos mitocondriales alternativos. Además, los codones AUA, AUC, AUU y son todos los codones de inicio permisibles.
| Organismo | Codón | Estándar | Novela |
|---|---|---|---|
| Mamífero | AGA, AGG | Arginina | Codón de parada |
| AUA | Isoleucina | La metionina | |
| UGA | Codón de parada | El triptófano | |
| Invertebrados | AGA, AGG | Arginina | Serina |
| AUA | Isoleucina | La metionina | |
| UGA | Codón de parada | El triptófano | |
| Levadura | AUA | Isoleucina | La metionina |
| UGA | Codón de parada | El triptófano | |
| CUA | La leucina | Treonina |
Algunas de estas diferencias deben ser considerados como pseudo-cambios en el código genético, debido al fenómeno de la La edición de ARN, que es común en las mitocondrias. En las plantas superiores, se pensó que codifica para CGG triptófano y no arginina; Sin embargo, el codón en el ARN procesado se descubrió que era el codón UGG, en consonancia con el código genético universal para el triptófano. Es de destacar que el código genético mitocondrial de artrópodos ha sufrido una evolución paralela dentro de un filo, con algunos organismos traducir únicamente AGG a la lisina.
Genomas mitocondriales tienen muchos menos genes que el eubacterias de la que se cree que son descendientes. Aunque algunos se han perdido por completo, muchos han sido transferidos a la núcleo, tales como las respiratorias complejo II subunidades de proteínas. Esto se cree que es relativamente común en el tiempo evolutivo. Unos pocos organismos, tales como la Cryptosporidium, en realidad tienen mitocondrias que carecen de cualquier ADN, presumiblemente debido a que todos sus genes se han perdido o transferidos. En Cryptosporidium, los mitocondrias tienen un alterada ATP sistema de generación que hace que el resistentes a muchos mitocondrial clásica parásito inhibidores tales como cianuro, azida, y atovacuona.
Replicación y herencia
Las mitocondrias se dividen por fisión binaria similar a la división celular bacteriana; a diferencia de las bacterias, sin embargo, las mitocondrias pueden también fundirse con otras mitocondrias .. La regulación de esta división se diferencia entre los eucariotas. En muchos eucariotas unicelulares, su crecimiento y división está vinculada a la ciclo celular. Por ejemplo, una sola mitocondria puede dividir de forma sincrónica con el núcleo. Este proceso de división y segregación debe ser estrechamente controlada de manera que cada célula hija reciba al menos una mitocondria. En otros eucariotas (en los seres humanos, por ejemplo), las mitocondrias pueden replicar su ADN y dividir principalmente en respuesta a las necesidades de energía de la célula, en lugar de en fase con el ciclo celular. Cuando las necesidades de energía de una célula son altos, las mitocondrias crecen y se dividen. Cuando el uso de energía es bajo, las mitocondrias se destruyen o se vuelven inactivos. En tales ejemplos, y en contraste con la situación en muchos eucariotas unicelulares, las mitocondrias son aparentemente distribuidos al azar a las células hijas durante la división de la citoplasma.
Genes mitocondriales de un individuo no son heredados por el mismo mecanismo que los genes nucleares. En la fecundación de un óvulo por un espermatozoide, el núcleo de huevo y cada núcleo del espermatozoide contribuyen igualmente a la composición genética de la núcleo del cigoto. En contraste, la mitocondria, y por lo tanto el ADN mitocondrial, por lo general viene de sólo el huevo. Las mitocondrias de los espermatozoides entren en el huevo, pero no contribuye a la información genética del embrión. En lugar de ello, las mitocondrias paternos están marcados con ubiquitina para seleccionar para su destrucción posterior dentro de la embrión. La célula huevo contiene relativamente pocas mitocondrias, pero es estas mitocondrias que sobreviven y se dividen para poblar las células del organismo adulto. Las mitocondrias son, por lo tanto, en la mayoría de los casos heredados por la línea femenina, conocida como herencia materna. Este modo se ve en la mayoría de los organismos, incluyendo todos los animales. Sin embargo, las mitocondrias en algunas especies a veces se pueden heredar paternalmente. Esta es la norma entre cierta plantas coníferas, aunque no en los árboles de pino y tejos. También se ha sugerido que se produce a un nivel muy bajo en los seres humanos.
Herencia uniparental conduce a poca oportunidad para la recombinación genética entre diferentes linajes de las mitocondrias, aunque una sola mitocondria puede contener 2-10 copias de su ADN. Por esta razón, generalmente se cree que el ADN mitocondrial de reproducir por fisión binaria. Lo recombinación no tener lugar mantiene la integridad genética en lugar de mantener la diversidad. Sin embargo, hay estudios que muestran evidencia de recombinación en el ADN mitocondrial. Es evidente que las enzimas necesarias para la recombinación están presentes en células de mamífero. Además, la evidencia sugiere que las mitocondrias de los animales pueden sufrir recombinación. Los datos son un poco más controvertida en los seres humanos, aunque existe evidencia indirecta de la recombinación. Si no se produce la recombinación, toda la secuencia de ADN mitocondrial representa un solo haplotipo, que lo hace útil para el estudio de la historia evolutiva de las poblaciones.
Estudios de genética de poblaciones
La casi ausencia de recombinación genética en el ADN mitocondrial hace que sea una fuente de información útil para los científicos que participan en genética de poblaciones y biología evolutiva . Debido a que todo el ADN mitocondrial se hereda como una sola unidad, o haplotipo, las relaciones entre el ADN mitocondrial de diferentes individuos pueden ser representados como una árbol de genes. Patrones en estos árboles de genes pueden ser utilizados para inferir la historia evolutiva de las poblaciones. El ejemplo clásico de esto es en genética evolutiva humanos, donde el reloj molecular se puede utilizar para proporcionar una fecha reciente para Eva mitocondrial . Esto se interpreta a menudo como un fuerte apoyo a una reciente expansión humana moderna fuera de Africa. Otro ejemplo humano es la secuencia de ADN mitocondrial de Neanderthal huesos. El relativamente a gran distancia evolutiva entre las secuencias de ADN mitocondrial de neandertales y los humanos que viven se ha interpretado como una prueba de la falta de cruce entre neandertales y los humanos anatómicamente modernos.
Sin embargo, el ADN mitocondrial refleja la historia de sólo las hembras en una población y por lo tanto puede no representar la historia de la población como un todo. Esto puede ser parcialmente superada por el uso de secuencias genéticas paternos, como la región de la no recombinación Y-cromosoma. En un sentido más amplio, sólo los estudios que incluyen también ADN nuclear puede proporcionar una historia evolutiva integral de una población.
Disfunción y la enfermedad
Las enfermedades mitocondriales
Con su lugar central en el metabolismo celular, el daño - y disfunción posterior - en las mitocondrias es un factor importante en una amplia gama de enfermedades humanas. Los trastornos mitocondriales a menudo presentes como trastornos neurológicos, pero puede manifestarse como miopatía, diabetes , endocrinopatía múltiple, o una variedad de otras manifestaciones sistémicas. Enfermedades causadas por mutación en el ADN mitocondrial incluyen Síndrome de Kearns-Sayre, Síndrome MELAS y La neuropatía óptica hereditaria de Leber. En la gran mayoría de los casos, estas enfermedades se transmiten por una hembra a sus hijos, como la cigoto deriva sus mitocondrias y por lo tanto su ADN mitocondrial del óvulo. Enfermedades tales como Síndrome de Kearns-Sayre, síndrome de Pearson, y oftalmoplejía externa progresiva se cree que es debido a reordenamientos de ADNmt a gran escala, mientras que otras enfermedades tales como Síndrome MELAS, La neuropatía óptica hereditaria de Leber, la epilepsia mioclónica con fibras rojas rasgadas (MERRF), y otros se deben a señalar mutaciones en el ADNmt.
En otras enfermedades, defectos en genes nucleares conducen a la disfunción de las proteínas mitocondriales. Este es el caso en La ataxia de Friedreich, paraplejia espástica hereditaria, y Enfermedad de Wilson. Estas enfermedades se heredan en un relación de dominación, como se aplica a la mayoría de otras enfermedades genéticas. Una variedad de trastornos puede ser causada por mutaciones nucleares de enzimas fosforilación oxidativa, tales como La deficiencia de coenzima Q10 y Síndrome de Barth. Las influencias ambientales pueden también interactuar con predisposiciones hereditarias y causar la enfermedad mitocondrial. Por ejemplo, puede haber un vínculo entre exposición a los pesticidas y la aparición posterior de Enfermedad de Parkinson.
Otras enfermedades que no están directamente vinculados a las enzimas mitocondriales pueden ofrecer la disfunción de las mitocondrias. Estos incluyen la esquizofrenia , trastorno bipolar, demencia, enfermedad de Alzheimer , la enfermedad, de Parkinson epilepsia , accidente cerebrovascular , enfermedad cardiovascular, retinitis pigmentosa, y diabetes mellitus . El hilo común que une a estas condiciones aparentemente no relacionados es el daño celular que causa el estrés oxidativo y la acumulación de especies reactivas al oxigeno. Estos oxidantes entonces dañan el ADN mitocondrial, lo que resulta en la disfunción mitocondrial y la muerte celular.
Las posibles relaciones con el envejecimiento
Dado el papel de las mitocondrias como potencia de la célula, puede haber alguna fuga de los de alta energía de electrones en la cadena respiratoria para formar especies reactivas al oxigeno. Esto puede resultar en significativos estrés oxidativo en las mitocondrias con altas tasas de mutación del ADN mitocondrial. Un círculo vicioso se cree que ocurre, ya que el estrés oxidativo conduce a mutaciones del ADN mitocondrial, que pueden conducir a anormalidades enzimáticas y más estrés oxidativo. Una serie de cambios se producen a las mitocondrias durante el proceso de envejecimiento. Los tejidos de pacientes de edad avanzada muestran una disminución de la actividad enzimática de las proteínas de la cadena respiratoria. Las grandes deleciones en el genoma mitocondrial pueden conducir a altos niveles de el estrés oxidativo y la muerte neuronal en Enfermedad de Parkinson. Enlaces hipotética entre el envejecimiento y el estrés oxidativo no son nuevos y se propusieron hace más de 50 años; Sin embargo, hay mucho debate sobre si los cambios mitocondriales son causas del envejecimiento o simplemente características de envejecimiento. Un estudio notable en los ratones no demostró aumento de las especies reactivas de oxígeno pesar de la creciente mutaciones del ADN mitocondrial, lo que sugiere que el proceso de envejecimiento no se debe a estrés oxidativo. Como resultado, las relaciones exactas entre las mitocondrias, el estrés oxidativo y el envejecimiento aún no se han resuelto.