

Massospondylus
Renseignements généraux
SOS Enfants a essayé de rendre le contenu plus accessible Wikipedia par cette sélection des écoles. Voulez-vous savoir sur le parrainage? Voir www.sponsorachild.org.uk
| Massospondylus Plage temporelle: Jurassique inférieur, 200-183Ma | |
|---|---|
 | |
| Vue d'artiste du Massospondylus représente l'animal bipède | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Classe: | Reptilia |
| Ordre: | Saurischia |
| Clade: | † Sauropodomorpha |
| Clade: | † Plateosauria |
| Clade: | † Massopoda |
| Famille: | † Massospondylidae |
| Genre: | † Massospondylus Owen, 1854 |
| Espèce | |
| Synonymes | |
| |
Massospondylus (pron .: / ˌ m æ s ɵ s p ɒ n ré ɨ l ə s / mas-OH- SPON di-ləs; du grec , μάσσων (Masson, "plus") et σπόνδυλος (spondylos, "vertèbres")) est un genre de prosauropode dinosaures du début du Jurassique Période ( Hettangien à Pliensbachien âges, ca. 200-183 Il ya des millions d'années). Il a été décrit par Sir Richard Owen en 1854 à partir de restes trouvés dans l'Afrique du Sud , et est donc l'un des premiers dinosaures d'avoir été nommé. Les fossiles ont été trouvés depuis à d'autres endroits en Afrique du Sud, le Lesotho et le Zimbabwe . Matériel de Arizona Formation Kayenta, l'Inde et l'Argentine a été affecté à ce genre à différents moments, mais le Arizonan et matérielle argentine sont maintenant affectés à d'autres genres.
Le Type espèce est M. carinatus; sept autres espèces ont été nommés au cours des 150 dernières années, mais seulement M. kaalae parmi ceux-ci est toujours considéré comme valide. Prosauropode systématique ont subi de nombreuses révisions au cours des dernières années, et de nombreux scientifiques en désaccord où exactement Massospondylus se trouve sur l'arbre évolutif des dinosaures. Le nom de famille Massospondylidae était autrefois inventé pour le genre, mais parce que la connaissance des relations prosauropode est dans un état de flux, il est difficile de savoir quels autres dinosaures-le cas échéant-appartiennent à un groupement naturel de massospondylids; plusieurs articles 2007 soutiennent la validité de la famille.
Bien Massospondylus a longtemps été décrite comme quadrupède, une étude de 2007 a trouvé pour être bipède. Ce était probablement un mangeur de plantes ( herbivores ), mais il est supposé que les prosauropodes ont peut-être été omnivores. Cet animal, 4-6 mètres (13-20 pi) de long, avait un long cou et de la queue, avec une petite tête et le corps mince. Sur chacune de ses pattes de devant, il portait une griffe de pouce forte qui a été utilisé dans la défense ou l'alimentation. Des études récentes indiquent Massospondylus n'a cessé de croître tout au long de sa durée de vie, possédé sacs similaires à ceux des oiseaux de l'air, et ont peut-être pris en charge pour ses jeunes.
Description

Massospondylus était un de taille moyenne prosauropode qui était autour de 4 mètres (13 pi) de longueur et pesait environ 135 kg (300 lb), bien que quelques sources ont estimé sa longueur jusqu'à 6 mètres (20 pi). Ce était un prosauropode typique avec un corps mince, un long cou et proportionnellement très petite tête. La colonne vertébrale est composée de neuf cervical (cou) vertèbres, 13 dorsale (dos) vertèbres, trois sacrée (hanche) vertèbres, et au moins 40 caudale (queue) vertèbres. Le pubis face avant, comme avec la plupart saurischiens . Il y avait une construction plus légère que celle de Plateosaurus , un dinosaure prosauropode ailleurs similaire. Le cou était proportionnellement plus que dans la plupart des autres prosauropodes, avec les plus grands cervicales étant quatre fois plus aussi large. Les membres antérieurs étaient seulement la moitié de la longueur des membres postérieurs mais très puissant, comme indiqué par la large extrémité supérieure de la humérus qui fourni zones de fixation pour une grande musculature du bras. Comme Plateosaurus, il avait cinq doigts à chaque main et du pied. La main était court et large, avec une grande faucille pouce en forme de griffe utilisé pour l'alimentation ou à la défense contre les prédateurs. Le pouce est le doigt le plus long dans la main, tandis que les quatrième et cinquième chiffres étaient minuscules, donnant les pattes de devant un regard en coin.
Anatomie crânienne
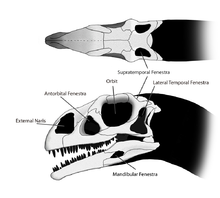
La petite tête de Massospondylus était d'environ la moitié de la longueur de la fémur. De nombreuses ouvertures, ou fenestrae, dans le crâne réduit son poids et a fourni un espace pour la fixation des muscles et des organes sensoriels. Ces fenestrae étaient présentes par paires, un de chaque côté du crâne. A l'avant du crâne avait deux grandes, elliptiques narines, qui étaient à peu près la moitié de la taille des orbites. Les orbites étaient proportionnellement plus important dans Massospondylus que dans genres apparentés tels que Plateosaurus. Le antéorbitaire fenestrae, plus petit que ceux observés dans Plateosaurus, était situé entre les yeux et le nez. A l'arrière du crâne avait deux autres paires de fenestrae temporelle: la fenestrae latéral temporal immédiatement derrière les orbites, qui ont été en forme de "T" inversé dans Massospondylus et le fenestrae supratemporal sur le dessus du crâne. Petit fenestrae également pénétré chaque mandibule. La forme du crâne est traditionnellement restauré comme plus large et plus courte que celle de Plateosaurus, mais cet aspect peut être due à une différence de seulement écrasement subi par les différents échantillons. Certaines fonctionnalités du crâne sont variables entre les individus; par exemple, l'épaisseur de la bordure supérieure de l'orbite et la hauteur de la partie postérieure maxillaire. Ces différences peuvent être dues à dimorphisme sexuel ou la variation individuelle.

nombre de dents est variable entre les individus et augmente avec la taille du crâne. Le prémaxillaire montre un nombre constant de 4 dents de chaque côté dans tous les crânes connus, mais dans le maxillaire gammes de nombre de dents de 14 à 22. Il existe 26 dents de chaque côté de la mâchoire inférieure dans le crâne connu plus grand. La hauteur des couronnes de dents diminue d'avant en arrière de la mâchoire supérieure mais était plus ou moins constante dans la mâchoire inférieure. L'absence d'usure des dents prononcée et la hauteur variable des couronnes suggère que les dents ont été remplacés par de nouveaux succédant à des intervalles de temps relativement courts. En particulier, il y avait variation de morphologie dentaire sur la base de la position des dents dans la mâchoire. Le heterodonty présent dans Massospondylus est supérieure à celle présente dans Plateosaurus , bien que sans surprise pas aussi prononcée que la spécialisation des dents Heterodontosaurus. Les dents plus étroites à l'avant du museau ont des sections transversales rondes et à des points effilés, contrairement aux dents du fond, qui sont spatulées et avait sections ovales.
Comme avec d'autres prosauropodes, il a été proposé que Massospondylus avaient joues. Cette théorie a été proposé parce qu'il ya quelques grands trous pour les vaisseaux sanguins sur les surfaces des os de la mâchoire, à la différence des nombreux petits trous présents sur les mâchoires de reptiles sans carcasse. Les joues auraient empêché la nourriture de se répandre quand Massospondylus mangé. Crompton et Attridge (1986) ont décrit des crânes Massospondylus comme possédant prononcée supraclusies et ont suggéré la présence d'un bec corné à la pointe de la mâchoire inférieure pour compenser la différence de longueur et de tenir compte de l'usure des dents sur les dents à l'extrémité du museau. Cependant, la différence de longueur peut être une erreur d'interprétation sur la base de broyage dans un plan de haut en bas, et la possession d'un bec est considéré comme improbable dans des études récentes.
Découverte
Les premiers fossiles de Massospondylus ont été décrites par le paléontologue Sir Richard Owen en 1854. À l'origine, Owen ne reconnaissait pas ces trouvailles que ceux d'un dinosaure; au contraire, il les attribue à des «grandes, éteint, reptiles carnivores» qui ont été liés à des lézards, des caméléons et des iguanes d'aujourd'hui. Ce matériau, une collection de 56 os, a été trouvé en 1853 par l'arpenteur du gouvernement Joseph Millard Orpen dans le Formation supérieure à Elliot Harrismith, Afrique du Sud et a été remis à la Musée Hunterian au Collège royal des chirurgiens à Londres. Parmi les vestiges étaient vertèbres du cou, le dos et la queue; une lame de l'épaule; une humérus; un bassin partielles; un fémur; une tibia; et les os des mains et des pieds. Tous ces os ont été trouvés désarticulé, ce qui rend difficile de déterminer si tout le matériel appartient à la même espèce ou non. Cependant, Owen a pu distinguer trois types de vertèbres caudales, qu'il attribue à trois genres différents différents:. Pachyspondylus, Leptospondylus et Massospondylus Massospondylus est séparé des deux autres genres sur la base de son beaucoup plus caudale vertèbres, qui entraînent également le nom scientifique qui a été dérivée des grecs termes Masson / μάσσων 'plus' et spondylos / σπόνδυλος «vertèbre». Cependant, il a été montré plus tard que les vertèbres caudales putatif de Massospondylus étaient vertèbres cervicales et fait que tout le matériel appartient probablement seulement une espèce de singe. Le 10 mai 1941, le Musée Hunterian a été démolie par une bombe allemande, détruisant tous les fossiles; seulement moulages restent.
Massospondylus possibles restes ont été retrouvés dans la Formation de Haute Elliot, Formation Clarens, et de la Bushveld Grès de l'Afrique du Sud et le Lesotho ; la Forêt Grès et de la Karroo supérieur Grès du Zimbabwe ; et le Formation des Kayenta Arizona. Ces restes sont constitués d'au moins 80 squelettes partiels et quatre crânes, représentant à la fois jeunes et les adultes. Le rapport de Massospondylus de la Formation Kayenta Arizona est basé sur un crâne décrit en 1985. Le crâne du spécimen Kayenta de l'Arizona est 25% plus grand que le plus grand crâne de tout spécimen africain. Le spécimen Kayenta possède quatre dents dans la prémaxillaire et seize dans le maxillaire. Unique parmi les dinosaures, il y avait aussi minuscule, un millimétriques (0,04 in-) dents longues palatine. Réétudier récente de crânes africaine Massospondylus, cependant, indique que le spécimen Kayenta ne concernent pas Massospondylus. Ce crâne Kayenta et éléments postcrâniens associés, identifiés collectivement comme MCZ 8893, a été récemment visé au genre nouvellement décrit Sarahsaurus.
Massospondylus avait également été signalée en Argentine , mais cela a été réévaluée comme un genre étroitement liés mais distincts. Les fossiles inclus plusieurs squelettes partiels et au moins un crâne, trouvés dans le Jurassique inférieur Formation Canon del Colorado San Juan, en Argentine. Cette matière a été nommé Adeopapposaurus en 2009.
Espèce

De nombreuses espèces ont été nommées, bien que la plupart ne sont plus considérés comme valides. M. carinatus, nommé par Richard Owen, est le espèce type. Autres espèces citées comprennent: M. browni ( Seeley, 1895), M. harriesi ( Broom 1911), M. hislopi ( Lydekker, 1890), M. huenei (Cooper, 1981), M. kaalae (Barrett 2009), M. rawesi (Lydekker, 1890), et M. schwarzi ( Haughton, 1924).
M. browni, M. harriesi, et M. schwarzi ont tous été trouvés dans la Formation de Haute-Elliot Province du Cap, Afrique du Sud. Tous les trois sont basées sur le matériel fragmentaire, et étaient considérés comme des indéterminée dans l'étude la plus récente. M. browni est basée sur deux col de l'utérus, deux pas en arrière, et trois caudal vertèbres et des éléments des pattes postérieures divers. M. harriesi est connu d'un membre antérieur bien conservé et les pièces d'un membre postérieur. M. schwarzi est connu d'un membre postérieur incomplet et sacrum. M. hislopi et M. rawesi ont été nommés à partir de fossiles trouvés dans l'Inde . M. hislopi est basée sur les vertèbres de la Trias supérieur Formation des Maleri Andhra Pradesh, tandis que M. rawesi se appuie sur une dent de la Formation du Crétacé supérieur de Takli Maharashtra. M. hislopi a été provisoirement retenu comme sauropodomorph indéterminée dans le dernier examen, mais M. rawesi peut être un théropode ou nondinosaur. M. huenei est une combinaison dérivée par Cooper pour Lufengosaurus huenei, car il considérait Lufengosaurus et Massospondylus être synonymes. Cette synonymie ne est plus acceptée.
M. kaalae a été décrite en 2009 sur la base d'un crâne partiel de la Formation de la Haute-Elliot dans Cap oriental en Afrique du Sud. Cette espèce est connue à la même période et la région que certains spécimens de M. carinatus. Elle diffère de l'espèce type dans la morphologie de la boîte crânienne, ainsi que dans plusieurs autres caractères du crâne telles que les proportions de la prémaxillaire.
Noms douteuses
Plusieurs dinosaures sont souvent considérés comme synonyme de Massospondylus. Il se agit notamment de la Leptospondylus mentionné ci-dessus et Pachyspondylus ainsi que Aristosaurus, Dromicosaurus, Gryponyx taylori et Hortalotarsus, qui sont des noms douteux de peu de valeur scientifique. Hortalotarsus skirtopodus a été nommé par Harry Seeley en 1894. Selon Broom (1911), "A l'origine la plupart du squelette était dans la roche, et il a été considéré par les agriculteurs comme le squelette d'un Bushman, mais il est dit avoir été détruit par la crainte qu'un Bushman squelette dans la roche pourrait avoir tendance à affaiblir la croyance religieuse de la génération montante ". Certains os de la jambe partielles ont été récupérés. Avec Massospondylus carinatus, Owen nommé Leptospondylus capensis et Pachyspondylus orpenii. Aristosaurus erectus a été nommé par ECN van Hoepen en 1920 basée sur un squelette presque complet. Hoepen également appelé Dromicosaurus gracile, qui consistait en un squelette partiel. Gryponyx taylori a été nommé par Sidney H. Haughton en 1924. Il se compose d'os de la hanche. Tous les fossiles ci-dessus proviennent des stades fauniques Hettangien ou Sinémurien de l'Afrique du Sud, où Massospondylus a été trouvé. Selon les règles de nomenclature zoologique, ces noms sont synonymes juniors. Ils ont été nommés après Massospondylus a été décrit dans un article scientifique; le nom Massospondylus prend donc la priorité.
Ignavusaurus, connu d'un jeune spécimen, peut également être synonyme de Massospondylus.
Classification
|
Est un Massospondylus prosauropode, un regroupement de début saurischian dinosaures qui ont vécu durant le Trias et du Jurassique, mais qui avait disparu à la fin du Jurassique. Les autres membres du groupe comprennent Plateosaurus , Yunnanosaurus, et Riojasaurus. Systématique des sauropodomorph basales continuent à subir une révision, et de nombreux genres autrefois considérées comme prosauropodes classiques ont récemment été retiré du groupe en Classification phylogénétique, au motif que leur inclusion ne constituerait pas une clade (un regroupement naturel contenant tous les descendants d'un même ancêtre commun). Exactement quels animaux constituer une monophylétique (groupement naturel) de prosauropodes est pas claire. Yates et Kitching (2003) a publié un clade constitué de Riojasaurus, Plateosaurus, Coloradisaurus, Massospondylus, et Lufengosaurus. Galton et Upchurch (2004) inclus Ammosaurus , Anchisaurus, Azendohsaurus, Camelotia, Coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Mussaurus, Plateosaurus, Riojasaurus, Ruehleia, Saturnales, Sellosaurus , Thecodontosaurus, Yimenosaurus et Yunnanosaurus dans un Prosauropoda monophylétique. Wilson (2005) considéré comme Massospondylus, Jingshanosaurus, Plateosaurus et Lufengosaurus un groupe naturel, Blikanasaurus et Antetonitrus sauropodes possibles. Bonnan et Yates (2007) considérées Camelotia, Blikanasaurus et Melanorosaurus sauropodes possibles. Yates (2007) placé Antetonitrus, Melanorosaurus et Blikanasaurus que sauropodes basales et a refusé d'utiliser le terme de Prosauropoda, car il considérait qu'il synonyme de Plateosauridae. Cependant, il n'a pas exclu la possibilité qu'un petit groupe de prosauropodes consistant Plateosaurus, Riojasaurus, Massospondylus et leurs parents les plus proches étaient monophylétique.
Massospondylus est le Type genre de la famille proposée Massospondylidae, à laquelle il donne son nom. La famille Massospondylidae peut également inclure Yunnanosaurus, bien que Lu et al. (2007) placé Yunnanosaurus dans sa propre famille. Yates (2007) considéré comme Massospondylus, Coloradisaurus et massospondylids Lufengosaurus, avec en Yunnanosaurus Anchisauria. Smith et Pol (2007) ont également trouvé un Massospondylidae dans leur analyse phylogénétique, y compris Massospondylus, Coloradisaurus et Lufengosaurus, ainsi que leur nouveau genre, Glacialisaurus. Adeopapposaurus, basée sur les fossiles que l'on croyait appartenir à une forme sud-américaine de Massospondylus, a également été classé comme un massospondylid, comme ce était Leyesaurus, un autre genre Amérique du Sud qui a été nommé en 2011. Pradhania a été considéré comme un basale sauropodomorph mais la nouvelle analyse cladistique effectuée par Novas et al., 2011 suggère que Pradhania est un massospondylid. Pradhania présente deux traits communs de la Massospondylidae récupérés dans leur analyse phylogénétique, et les fossiles de Pradhania ont été découverts de la même région et lavabo en Inde comme M. hislopi.
Paléoécologie
Le faunes et flores du Jurassique étaient similaires dans le monde entier, avec conifères adaptés pour le temps chaud devenir les plantes communes, et prosauropodes et basales théropodes les constituants principaux de monde une faune de dinosaures. L'environnement du début du Jurassique Afrique australe a été décrit comme un désert. Afrique Massospondylus était un contemporain de Temnospondyli; tortues ; une Sphenodontia; Rauisuchia; tôt crocodylomorphs; tritylodontid et trithelodontid thérapsides; morganucodontid mammifères ; et les dinosaures, y compris le petit théropode Megapnosaurus rhodesiensis et plusieurs genres de début ornithischiens , comme Lesothosaurus et heterodontosaurids Abrictosaurus, Heterodontosaurus, Lycorhinus et Pegomastax. Jusqu'à récemment, Massospondylus était considéré comme le seul sauropodomorph connue de la Formation de la Haute-Elliot. Cependant, les découvertes plus récentes ont révélé une faune diversifiée de sauropodomorph contemporaines avec six autres espèces, y compris les prosauropodes Ignavusaurus, Arcusaurus et deux taxons sans nom ainsi que deux sauropodes anonymes.
Il ne est pas clair qui carnivores peuvent avoir sur la proie Massospondylus. La plupart des théropodes qui ont été découverts dans des roches d'âge Jurassique inférieur en Afrique australe, comme Megapnosaurus, étaient plus petits que prosauropodes moyennes comme Massospondylus. Ces petits prédateurs ont été postulé que l'utilisation attaques sabrer rapide à l'usure bas prosauropodes, qui aurait pu se défendirent avec leurs grandes griffes mains et les pieds. Le 6-mètre-(20) Bien immobilier à long théropode carnivore Dracovenator vécu pendant la même période (Hettangien à des stades Sinémurien) que Massospondylus et a également été trouvé dans la Formation Elliot d'Afrique du Sud.
Paléobiologie
Comme avec tous les dinosaures, une grande partie de la biologie de Massospondylus, y compris son comportement, la coloration et la physiologie, reste inconnu. Cependant, des études récentes ont permis à la spéculation informés sur des sujets tels que les modèles de croissance, l'alimentation, la posture, la reproduction et la respiration.
Une étude de 2007 a suggéré que Massospondylus peut avoir utilisé ses bras courts dans la défense contre les prédateurs (les «claques défensives»), dans intraspécifique combat ou dans l'alimentation, bien que ses bras étaient trop courts pour atteindre sa bouche. Les scientifiques spéculent que Massospondylus aurait pu utiliser sa grande pollex (pouce) griffe au combat, à dépouiller matériel végétal des arbres, creuser, ou pour le toilettage.
Croissance

Une étude de 2005 a indiqué que le taxon soeur de Massospondylus, Plateosaurus, expose des modèles de croissance affectés par des facteurs environnementaux. L'étude a indiqué que lorsque la nourriture était abondante, ou lorsque le climat était favorable, Plateosaurus affiché une croissance accélérée. Ce modèle de croissance est appelé «plasticité du développement". Il est invisible dans d'autres dinosaures, y compris Massospondylus, malgré la relation étroite entre Plateosaurus et Massospondylus. L'étude a indiqué que Massospondylus grandi le long d'une trajectoire de croissance spécifique avec peu de variation dans le taux de croissance et la taille finale d'un individu. Une autre étude de détermination de l'âge indiqué que Massospondylus ont augmenté à un taux maximal de 34,6 kg (76,3 lb) par an et a été continue de croître à environ 15 ans.
Régime
Prosauropodes tels que Massospondylus peuvent avoir été herbivores ou omnivores. Aussi récemment que les années 1980, les paléontologues ont débattu de la possibilité de carnivore dans prosauropodes. Toutefois, l'hypothèse d'prosauropodes carnivores a été discréditée, et toutes les études récentes de favoriser un mode de vie herbivores ou omnivores pour ces animaux. Galton et Upchurch (2004) ont constaté que les caractéristiques crâniennes (comme articulation de la mâchoire) de la plupart des prosauropodes sont plus proches de ceux des reptiles herbivores que celles de ceux carnivores, et la forme de la dent couronne est similaire à ceux des herbivores moderne ou omnivores iguanes. La largeur maximale de la couronne était supérieure à celle de la racine, résultant en une arête de coupe similaires à ceux des reptiles herbivores ou omnivores existantes. Barrett (2000) a proposé que prosauropodes complété leurs régimes herbivores avec des petites proies ou charognes. Gastrolithes (pierres de gésier) ont été trouvés en association avec trois fossiles Massospondylus de la Forêt-Grès en Zimbabwe , et avec un animal -comme Massospondylus de la Trias supérieur de Virginie. Jusqu'à récemment, les scientifiques croyaient que ces pierres ont fonctionné comme un moulin gastrique pour aider l'ingestion de matériel végétal, de compensation pour son incapacité à mâcher, comme ce est le cas dans de nombreux oiseaux modernes. Cependant, Ailes et Sander (2007) ont montré que la nature poli et l'abondance de ces pierres excluaient une utilisation comme un moulin gastrique efficace dans la plupart non théropodes dinosaures, y compris Massospondylus.
La démarche et l'amplitude des mouvements

Bien longtemps supposé avoir été quadrupède, une étude anatomique de la forelimbs 2007 a remis en question, faisant valoir que leur gamme limitée de mouvement interdit la marche quadrupède habituelle efficace. Ni les membres antérieurs pourraient balancer avant et à l'arrière d'une manière similaire aux membres postérieurs, ne pouvaient être tourné la main avec les surfaces palmaires orientés vers le bas. Cette incapacité à pronation la main est également soutenue par in situ trouvailles de bras articulés (encore connectés) qui montrent toujours les mains avec unrotated palmaire visages face de l'autre. L'étude a également exclu la possibilité de «coup-de-pied" et d'autres formes de locomotion qui ferait un locomotion efficace possible sans la nécessité de pronation la main. Bien que sa masse suggère une nature quadrupède, Massospondylus aurait été limitée à ses pattes arrière de locomotion.
Depuis la découverte de rudimentaire et non fonctionnel clavicules dans cératopsiens on a supposé que ces os de l'épaule ont été réduits dans tous les dinosaures qui ne ont pas vrai furculae. Robert Bakker (1987) a suggéré que cela aurait permis à la omoplates de se balancer avec les membres antérieurs de dinosaures quadrupèdes, augmentant leur longueur des membres antérieurs fonctionnelle. Ce aurait réduit l'écart de longueur entre membres antérieurs et postérieurs dans un Massospondylus quadrupède. Cependant, une découverte récente montre que Massospondylus possédaient bien développé clavicules qui se sont attachés dans un arrangement de furcula-like, agissant comme un fermoir entre les omoplates droite et gauche et en interdisant toute rotation de ces os. Cette découverte indique que la réduction de la clavicule est limitée à la ligne d'évolution conduisant aux cératopsiens. Il indique également que la furcula des oiseaux est dérivé d'clavicules.
Michael Cooper (1981) a noté que la zygapophyses des vertèbres du cou ont été inclinés, interdisant un déplacement horizontal significatif du col, de sorte que "par conséquent tout mouvement important dans cette direction doit avoir été effectuée par un changement de la position du corps entier». Cela a été contredit dans une étude récente, notant que seuls les cervicales basalmost montrent zygapophyses inclinés, permettant un mouvement horizontal suffisante du cou comme un ensemble.
Reproduction

En 1976, un embrayage de sept 190 millions d'années-vieux oeufs Massospondylus ont été trouvés dans le parc national du Golden Gate Highlands en Afrique du Sud par James Kitching, qui les a identifiés comme appartenant plus susceptibles de Massospondylus. Il était près de 30 ans avant l'extraction a commencé sur les fossiles de la 15-centimétrique (y 6) longues embryons. Ils restent les plus anciens embryons de dinosaures jamais découverts. Au début de 2012, au moins 10 couvées d'au moins quatre horizons fossilifères avaient été trouvés, avec jusqu'à 34 oeufs par couvée. Cela indique que ce site de nidification peut avoir été utilisé à plusieurs reprises (fidélité au site), par groupes d'animaux (de nidification coloniale); dans les deux cas, ceux-ci représentent la preuve la plus ancienne de ce comportement. Structures sédimentaires indiquent que l'aire de nidification se trouvait à proximité d'un lac. Les coquilles d'œufs étaient très minces (environ 0,1 mm), permettant l'échange de gaz, même dans une faible teneur en oxygène et le dioxyde de carbone environnement riche, ce qui indique que les oeufs ont été au moins partiellement enterrés dans le substrat. Il n'y a pas de conseils qui Massospondylus nids construits; Cependant, la disposition des oeufs en rangs serrés indique que les œufs ont été poussés dans cette position par les adultes.
Les embryons représentaient probablement près de-nouveau-nés. Bien que les caractéristiques squelettiques étaient similaires à ceux des adultes, les proportions du corps étaient très dissemblables. La tête était grande avec un museau court et très grandes orbites, dont le diamètre se élève à 39% de la totalité de la longueur du crâne. Le cou est court, contrastant avec le très long cou dans les adultes. Os et caudales Ceinturon étaient relativement petit. Les membres antérieurs étaient de longueur égale aux membres postérieurs, ce qui indique que Massospondylus nouvellement éclos étaient quadrupède, contrairement aux adultes bipèdes. La découverte de l'éclosion empreintes avec des impressions manus confirmé leur quadrupedality. Ces impressions montrent que la main n'a pas été en pronation, avec la paume face à face de l'autre et le pouce vers l'avant. Le manus unpronated et la grosse tête indiquent qu'une locomotion efficace ne était pas possible pour nouvellement éclos Massospondylus. Notamment, les quasi-hachures ne avaient pas de dents, ce qui suggère qu'ils ne avaient aucun moyen de se nourrir. Sur la base de l'absence de dents et de la locomotion inefficaces, les scientifiques pensent que les soins postnatals aurait été nécessaire. Ce est en outre soutenue par des preuves que les hachures sont restés sur les sites de nidification jusqu'à ce qu'ils aient doublé de taille.

Nouvellement éclos mineurs sont connus d'une seconde prosauropode, Mussaurus; ces vestiges ressemblent à ceux de la Massospondylus embryonnaire, ce qui suggère que quadrupedality était présent dans nouvellement éclos Mussaurus et probablement d'autres prosauropodes ainsi. Le quadrupedality des hachures suggère que la posture quadrupède de suite sauropodes peuvent avoir évolué de la rétention des caractéristiques juvéniles chez les animaux adultes, un phénomène connu sous le nom de l'évolution Pédomorphose. Cette découverte "jette un peu de lumière dans les voies évolutives à travers lequel les adaptations particulières de dinosaures géants ont été atteints" donc, a déclaré le paléontologue français Eric Buffetaut.
Système respiratoire
Beaucoup saurischian dinosaures possédaient vertèbres et des côtes qui contenaient cavités creusées (pneumatique foramina), qui a réduit le poids des os et peut-être servi comme un système de base »de ventilation accréditive» similaire à celle des oiseaux modernes. Dans un tel système, les vertèbres du cou et des nervures sont creusées par le sac d'air col de l'utérus; les vertèbres du haut du dos, par le poumon; et le bas du dos et sacral (hanche) vertèbres, par le sac d'air abdominale. Ces organes constituent une méthode complexe et très efficace de la respiration. Prosauropodes sont le seul groupe important de saurischiens sans un vaste système de foramen pneumatique. Bien que possibles indentations pneumatiques ont été trouvés dans Plateosaurus et Thecodontosaurus, les empreintes étaient très petites. Une étude réalisée en 2007 a conclu que prosauropodes comme Massospondylus ont probablement eu sacs aériens abdominaux et cervicales, fondée sur la preuve pour eux dans soeur taxons (théropodes et sauropodes). L'étude a conclu qu'il était impossible de déterminer si prosauropodes avaient un poumon accréditives comme un oiseau, mais que les sacs d'air sont presque certainement présents.