

Cervelet
Renseignements généraux
Enfants SOS ont produit une sélection d'articles de wikipedia pour les écoles depuis 2005. Avant de vous décider à propos de parrainer un enfant, pourquoi ne pas en apprendre davantage sur différents organismes de parrainage premiers ?
| Cerveau: Cerebellum | ||
|---|---|---|
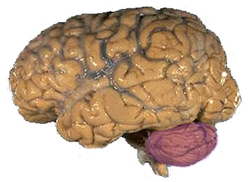 | ||
| Figure 1a: A cerveau humain, avec le cervelet en violet. | ||
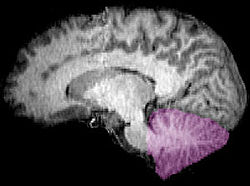 | ||
| Figure 1b: Image IRM montrant une mi- vue sagittale du cerveau humain, avec le cervelet en violet. | ||
| Partie de | Cerveau | |
| Artère | SCA, AICA, PICA | |
| Veine | supérieure, inférieur | |
Le cervelet est une région du cerveau qui jouent un rôle important dans l'intégration de la perception sensorielle, la coordination et commande du moteur. Afin de coordonner le contrôle moteur, il ya beaucoup de voies neuronales reliant le cervelet avec le cérébral cortex moteur (qui envoie des informations à la muscles provoquant leur déplacer) et le tractus spino-cérébelleuse (qui fournit proprioceptive rétroaction sur la position du corps dans l'espace). Le cervelet intègre ces voies, comme un conducteur de train, en utilisant la rétroaction constante sur la position du corps à des mouvements de moteur affiner.
En raison de cette fonction de «mise à jour» du cervelet, lésions son sein ne sont pas si débilitante à provoquer paralysie, mais plutôt présente comme déficits de rétroaction résultant des troubles dans un léger mouvement, équilibre, posture, et l'apprentissage moteur. Les premières observations par physiologistes au cours du 18ème siècle ont indiqué que les patients atteints de dommages cérébelleux montrent des problèmes avec la coordination motrice et de mouvement. La recherche sur la fonction cérébelleuse du début au milieu du 19ème siècle a été fait par des études de lésions et d'ablation en animaux . physiologistes de recherche ont noté que de telles lésions ont conduit à des animaux avec des mouvements étranges, démarche maladroite, et une faiblesse musculaire. Ces observations et les études ont conduit à la conclusion que le cervelet est une structure de commande de moteur. Cependant, la recherche moderne montre que le cervelet joue un rôle plus large dans un certain nombre de fonctions cognitives clés, y compris attention et le traitement de la langue , la musique , et d'autres stimuli sensoriels temporelles.
Caractéristiques générales
Le cervelet est situé dans la partie postérieure inférieure de la tête (le cerveau postérieur), directement à la dorsale pons, et inférieure à la lobe occipital (Figs. 1 et 3). En raison de son grand nombre de minuscules cellules granulaires, le cervelet contient plus de 50% de l'ensemble neurones dans le cerveau, mais il ne prend en hausse de 10% du volume total du cerveau. Le cervelet reçoit près de 200 millions de fibres d'entrée; en revanche, le nerf optique est composé de seulement un million de fibres.
Le cervelet est divisé en deux grandes hémisphères , tout comme le Cerebrum, et contient dix petits lobules. Le cytoarchitecture ( cellulaire organisation) du cervelet est très uniforme, avec des connexions organisées dans un rugueux, tridimensionnel perpendiculaire gamme de des éléments de circuit. Cette uniformité organisationnelle rend les circuits nerveux relativement facile à étudier. Pour imaginer ce «tableau perpendiculaire," on pourrait imaginer une rue bordée d'arbres avec des fils en allant tout droit à travers les branches d'un arbre à l'autre.
Développement et évolution
Au cours des premiers stades de le développement embryonnaire, le cerveau commence à se former en trois segments distincts: la prosencéphale, mésencéphale, et rhombencéphale. Le rhombencéphale est la plus caudale (vers la queue) segment du cerveau embryonnaire; ce est de ce segment que le cervelet se développe. Le long du segment rhombencéphalique embryonnaire développer huit gonflements, appelé rhombomères. Le cervelet découle de deux situés dans le rhombomères plaque alaire de la tube neural, une structure qui forme éventuellement le cerveau et la moelle épinière. Les rhombomères spécifiques à partir de laquelle les formes du cervelet sont rhombomère 1 (Rh.1) caudale (près de la queue) et l '"isthme" rostralement (près de l'avant).
Deux régions primaires sont censés donner naissance à des neurones qui constituent le cervelet. La première région est la zone ventriculaire dans le toit de la quatrième ventricule. Cette région produit Les cellules de Purkinje du cervelet et profonde neurones nucléaires. Ces cellules sont des neurones de sortie primaires du cortex cérébral et du cervelet. La deuxième zone germinative (berceau cellulaire) est connu comme la lèvre rhombique, les neurones puis passer par semaine embryonnaire 27 à la couche granulaire externe. Cette couche de cellules-trouve à l'extérieur du cervelet-produit les neurones granulaires. Les neurones granulaires migrent de cette couche extérieure pour former une couche interne dite couche interne de granules. La couche granulaire externe cesse d'exister dans le cervelet adulte, ne laissant que des cellules granulaires dans la couche granulaire interne. Le cervelet matière blanche peut être une troisième zone germinale dans le cervelet; Cependant, sa fonction de zone germinative est controversée.
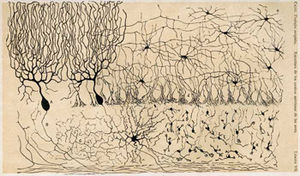
Le cervelet est de archipalliar origine phylogénétique. Le pallium est un terme pour la matière grise qui forme le cortex. Le Archipallium est l'un des plus évolutif régions cérébrales primitives. Les circuits du cortex cérébelleux ressemblent dans l'ensemble classes de vertébrés , y compris poissons , reptiles , oiseaux et mammifères (par exemple, Fig. 2). Cela a été considéré comme une preuve que le cervelet exerce des fonctions importantes à tous les vertébrés espèces .
Anatomie
Le cervelet contient similaire divisions de matière grise et blanche comme la cerveau. Embarqué dans la matière blanche, qui est connu comme le Arbor Vitae (Tree of Life) dans le cervelet en raison de ses ramifiés, arborescentes apparence sont quatre noyaux cérébelleux profonds. Trois segments phylogénétiques bruts sont largement regroupés par fonction générale. Les trois couches corticales contiennent différents types cellulaires qui créent souvent différentes boucles asservies et prédictives. oxygénée sang est fourni par trois branches artérielles au large de la basilaire et artères vertébrales.
Divisions
Le cervelet peut être divisé selon trois critères différents: anatomiques brute, phyologenetical et fonctionnels.
Divisions anatomiques brutes
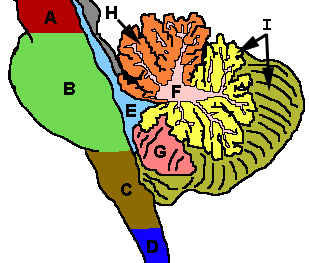
Lors de l'inspection grossière, trois lobes peuvent être distinguées dans le cervelet: le lobe flocculonodulaire, le lobe antérieur (rostral au «fissure primaire»), et le lobe postérieur (dorsal de la "fissure primaire»). Les deux derniers peuvent être subdivisés dans une ligne médiane vermis cérébelleux et hémisphères cérébelleux latérales.
 |  |
Divisions phylogénétiques et fonctionnels
Le cervelet peut également être divisée en trois parties sur la base à la fois critères phylogénétiques (l'âge de l'évolution de chaque partie) et sur des critères fonctionnels (les connexions entrantes et sortantes chaque partie a et le rôle joué dans la fonction cérébelleuse normale). De l'phylogénétiquement ancien au plus récent, les trois parties sont:
| Dénomination fonctionnelle (dénomination phylogénétique) | Parties anatomiques | Rôle |
| Vestibulocerebellum (archéocervelet) | Lobe flocculonodulaire (et vermis immédiatement adjacentes) | Le vestibulocerebellum régule l'équilibre et les mouvements oculaires. Il reçoit entrée vestibulaire à la fois du canaux semi-circulaires et de la noyaux vestibulaires, et envoie fibres revenir à la médiane et noyaux vestibulaires latéraux. Il reçoit également entrée visuelle à partir de la colliculus supérieur et de la cortex visuel (ce dernier via le noyaux pontiques, formant une voie cortico-ponto-cérébelleux). Les lésions de la cause de vestibulocerebellum troubles de l'équilibre et la marche. |
| Spinocerebellum (paléocervelet) | Vermis et des parties intermédiaires des hémisphères (les «paravermis") | Le spinocerebellum réglemente les mouvements du corps et des membres. Il reçoit entrée de la proprioception des colonnes dorsales de la la moelle épinière (y compris la tractus spino-cérébelleuse), ainsi que de la nerf trijumeau, ainsi que de visuel et systèmes auditifs. Il envoie des fibres à noyaux cérébelleux profonds projet qui à son tour à la fois au cortex cérébral et le tronc cérébral, fournissant ainsi de modulation diminuant les systèmes à moteur. Le spinocerebellum contient des cartes sensorielles comme il reçoit des données de la position de différentes parties du corps dans l'espace: en particulier, le vermis reçoit des fibres à partir du tronc et des parties proximales des branches, tandis que les parties intermédiaires des hémisphères reçoivent des fibres à partir des parties distales des branches . Le spinocerebellum est capable d'élaborer entrée proprioceptive afin d'anticiper la position future d'une partie du corps au cours d'un mouvement, d'une manière "feed forward". |
| Cérébro-cervelet (néocérébellum, Pontocerebellum) | Parties latérales de la hémisphères | Le néocérébellum est impliqué dans la planification de mouvement et évaluer l'information sensorielle pour l'action. Il reçoit une entrée exclusivement à partir du cortex cérébral (en particulier le lobe pariétal) via le noyau pontique (formant des voies cortico-ponto-cérébelleuse), et envoie des fibres essentiellement à la ventrolatérale thalamus (lui-même relié à des zones motrices du prémotrice cortex et aire motrice primaire du cortex cérébral) et à la noyau rouge (à son tour reliée à la noyau olivaire inférieur, ce qui nous ramène aux hémisphères cérébelleux). Le néocérébellum est impliqué dans le mouvement de planification qui est sur le point de se produire et a des fonctions purement cognitives ainsi. |
Une grande partie de ce qui est compris sur les fonctions du cervelet tiges de documentation minutieuse des effets des lésions focales chez des patients humains qui ont souffert de blessures ou de maladie ou par la recherche de la lésion des animaux.
Noyaux profonds
Les noyaux profonds de l'acte de cervelet que les principaux centres de communication, et les quatre noyaux différents du cervelet (denté, interposé, fastigial et vestibulaire) recevoir et envoyer des informations à des parties spécifiques du cerveau. En outre, ces noyaux recevoir les deux signaux excitateurs et inhibiteurs d'autres parties du cerveau qui à son tour affecte signaux sortants du noyau.
Couches corticales


Il existe trois couches au cortex cérébelleux; de l'extérieur vers l'intérieur, ce sont les couches moléculaires, Purkinje, et granulaires. La fonction du cortex cérébelleux est essentiellement à moduler l'information circulant à travers les noyaux profonds. Le microcircuits du cervelet est schématisé sur la figure 5. Mossy et fibres grimpantes portent des informations sensori-motrice dans les noyaux profonds, qui à son tour transmettre à divers domaines prémotrices, régulant ainsi la gagner et le calendrier des actions motrices. Mossy et des fibres grimpantes se nourrissent également cette information dans le cortex cérébelleux, qui effectue divers calculs, résultant dans la régulation des cellules de Purkinje tir. les neurones de Purkinje feed-back dans les noyaux profonds via un inhibitrice synapse. Cette synapse règle la mesure dans laquelle les fibres moussues et escalade activer les noyaux profonds et donc de contrôler l'effet ultime du cervelet sur la fonction motrice. La force synaptique de presque toutes les synapses dans le cortex cérébelleux a été montré pour subir plasticité synaptique. Cela permet à l'ensemble de circuits du cortex cérébelleux pour ajuster en continu et d'affiner la sortie du cervelet, formant la base de certains types d'apprentissage moteur et de coordination. Chaque couche dans le cortex cérébelleux contient les différents types de cellules qui composent ce circuit.
Couche granulaire
La couche la plus interne contient les corps cellulaires des deux types de cellules: les nombreux et minuscule cellules granulaires, ainsi que la plus grande Les cellules de Golgi. Les fibres moussues entrent dans la couche granulaire de leur principal point d'origine, les noyaux pontiques. Ces fibres forment des synapses excitatrices avec les cellules granulaires et des cellules des noyaux cérébelleux profonds. Les cellules granulaires envoient leurs axones connu en forme de T que fibres plan parallèle à la couche superficielle moléculaire, où ils forment des centaines de milliers de synapses avec cellule de Purkinje dendrites. Le cervelet humain contient de l'ordre de 60 à 80 milliards de cellules granulaires, ce qui rend ce type de cellule unique de loin les plus nombreux neurones dans le cerveau (environ 70% de tous les neurones dans le cerveau et la moelle épinière, combinés). cellules de Golgi fournissent une rétroaction inhibitrice de cellules granulaires, formant une synapse avec eux et en projetant un axone dans la couche moléculaire.
couche de Purkinje
La couche intermédiaire contient un seul type de corps, cette cellule de la grande Cellules de Purkinje. Les cellules de Purkinje sont les neurones d'intégration primaires du cortex cérébelleux et fournissent sa seule sortie. dendrites de cellules de Purkinje sont grandes tonnelles avec des centaines de branches épineuses atteignant dans la couche moléculaire (Fig. 6). Ces arbres dendritiques sont plates-la quasi-totalité d'entre eux se trouvent dans des plans-voisine avec tonnelles Purkinje dans des plans parallèles. Chaque fibre parallèle à partir des cellules granulaires exécute orthogonalement à travers ces tonnelles, comme un fil passant par de nombreuses couches. les neurones de Purkinje sont GABAergique-à-dire qu'ils ont synapses inhibitrices-avec les neurones du cervelet profonde et noyaux vestibulaires du tronc cérébral. Chaque cellule de Purkinje reçoit entrée excitatrice de 100 000 à 200 000 fibres parallèles. Fibres parallèles sont censés être responsables pour la simple (tout ou rien, amplitude invariant) fortification de la cellule de Purkinje.
Les cellules de Purkinje reçoivent également de l'entrée noyau olivaire inférieur via fibres grimpantes. Un bon moyen mnémotechnique pour cette interaction est l'expression «monter l'autre olivier", étant donné que les fibres proviennent de l'escalade olive inférieure controlatérale. En contraste frappant avec les entrées 100 000-Plus à partir de fibres parallèles, chaque cellule de Purkinje reçoit une entrée à partir de fibres exactement un d'escalade; mais cette fibre unique "monte" les dendrites de la cellule de Purkinje, enroulement autour d'eux et de faire un grand nombre de synapses comme il va. L'entrée nette est si forte qu'un seul potentiel d'action à partir d'une fibre de l'escalade est capable de produire un «pic complexe» dans la cellule de Purkinje: une rafale de plusieurs pointes dans une rangée, avec diminution d'amplitude, suivi d'une pause au cours de laquelle des pointes simples sont supprimées.
Couche moléculaire
Cette couche la plus externe du cortex cérébelleux contient deux types d'inhibiteur interneurones: la stellaire et cellules panier. Il contient également les arbres dendritiques de neurones de Purkinje et faisceaux de fibres parallèles des cellules granulaires. Les deux cellules étoilées et de basket forment synapses GABAergiques SUR DES dendrites des cellules de Purkinje.
Pédoncules
De même, le cervelet suit la tendance des «trois», avec trois grands pédoncules d'entrée et de sortie (faisceaux de fibres). Ce sont le supérieur (brachium conjunctivum), au milieu (brachium pontis), et inférieur (corps de restiforme) pédoncules cérébelleux.
| Pédoncule | Description |
| Supérieur | Bien qu'il existe des fibres afférentes de la tractus spino-cérébelleuse antérieure qui sont transmis au lobe cérébelleux antérieur par ce pédoncule, la plupart des fibres sont des fibres efférentes. Ainsi, le pédoncule cérébelleux supérieur est la voie de sortie majeure du cervelet. La plupart des fibres efférentes sont créés à l'intérieur de la noyau dentelé qui à son tour à divers projets y compris le mésencéphale structures noyau rouge, le noyau ventral latérale ventrale / antérieure de la thalamus, et le médullaire. Le dentatorubrothalamocortical (noyau dentelé> noyau rouge> thalamus> cortex prémoteur) et cerebellothalamocortical (cervelet> thalamus> cortex prémoteur) voies sont deux voies principales qui traversent cette pédoncule et qui sont importants dans la planification de moteur. |
| Milieu | Ce est entièrement composé de fibres afférentes originaires de la noyaux pontiques dans le cadre du massif voies corticopontocerebellar (cortex cérébral> pons> cervelet). Ces fibres descendent des aires sensorielles et motrices du cérébrale néocortex et faire le pédoncule cérébelleux moyen le plus grand des trois pédoncules cérébelleux. |
| Inférieur | Cela comporte plusieurs types de fibres d'entrée et de sortie qui sont principalement concernés par l'intégration entrée sensorielle proprioceptive avec moteur fonctions vestibulaires tels que l'équilibre et l'entretien de la posture. Informations proprioceptives du corps est effectué au cervelet via l'dorsale tractus spino-cérébelleuse. Ce tube passe à travers le pédoncule cérébelleux inférieur et des synapses dans le paléocervelet. Informations vestibulaire projette sur l'archéocervelet. Le fibres escalade de la inférieure terme d'olive à travers le pédoncule cérébelleux inférieur. Ce pédoncule porte également des informations directement à partir du Les cellules de Purkinje sur la noyaux vestibulaires du tronc cérébral dorsale située à la jonction entre la Pons et médullaire. |
Il ya trois sources d'entrée au cervelet, dans deux catégories constituées de fibres moussues et escalade, respectivement. Les fibres moussues peuvent provenir des noyaux pontiques, qui sont des groupes de neurones situés dans le pont qui transportent l'information du cortex cérébral controlatéral. Ils peuvent également se poser dans le tractus spino-cérébelleuse dont l'origine est située dans le ipsilatéral la moelle épinière. La plupart de la sortie provenant du cervelet synapses initialement sur les noyaux cérébelleux profonds avant de sortir par l'intermédiaire des trois pédoncules. L'exception la plus notable est l'inhibition directe des noyaux vestibulaires par des cellules de Purkinje.
L'approvisionnement en sang
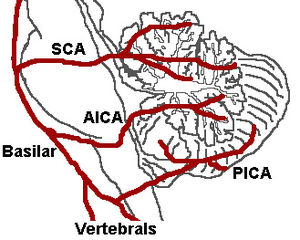
Trois artères approvisionnement en sang au cervelet (figure 7.): Le artère cérébelleuse supérieure (SCA), artère cérébelleuse antérieure inférieure (AICA), et Artère cérébelleuse postéro-inférieure (PICA).
Artère cérébelleuse supérieure
Les branches SCA large de la partie latérale de l'artère basilaire, juste inférieure à sa bifurcation dans l'artère cérébrale postérieure. Ici, il se enroule autour de la protubérance arrière (à laquelle elle fournit aussi du sang) avant d'atteindre le cervelet. Le SCA fournit le sang à la plupart du cortex cérébelleux, les noyaux cérébelleux, et les pédoncules cérébelleux moyens et supérieurs.
Artère cérébelleuse antéro-inférieure
Les branches de l'AICA au large de la partie latérale de l'artère basilaire, juste au-dessus de la jonction des artères vertébrales. Dès son origine, elle se ramifie le long de la partie inférieure de la protubérance à l'angle ponto-cérébelleux avant d'atteindre le cervelet. Cette artère fournit le sang à la partie antérieure inférieure du cervelet, et de la facial (NC VII) et nerfs vestibulaire (CN VIII).
Obstruction de l'AICA peut causer parésie, la paralysie, et la perte de sensation dans le visage; il peut aussi causer des troubles auditifs . En outre, il pourrait causer un infarctus de l'angle ponto-cérébelleux. Cela pourrait conduire à hyperacusia (dysfonctionnement du muscle de l'étrier, innervé par CN VII) et vertiges (mauvaise interprétation de la vestibulaire canal semi-circulaire de accélération endolymphe causé par altération de CN VIII).
Artère cérébelleuse postéro-inférieure
Les branches PICA large de la partie latérale des artères vertébrales juste inférieures à leur point de jonction avec l'artère basilaire. Avant d'arriver à la surface inférieure du cervelet, le PICA envoie branches dans le bulbe, fournir du sang à plusieurs noyaux des nerfs crâniens. Dans le cervelet, le PICA fournit le sang à la partie inférieure postérieure du cervelet, le pédoncule cérébelleux inférieur, le ambiguus noyau, le vague noyau moteur, la moelle noyau trijumeau, la noyau solitaire, et de la noyaux vestibulaire.
Fonction générale
Fonctionnellement, le fibres d'escalade et de la moussues voies de fibres cellulaires parallèle fibres granules sont les deux principaux types d'afférences au cervelet dans son ensemble et aux cellules de Purkinje en particulier. Ces systèmes diffèrent considérablement afférentes dans leur connectivité. La cellule de Purkinje et sa fibre afférente d'escalade ont une relation un-à-un et la projection globale est organisé pour produire une activation synchrone de groupes spécifiques de cellules de Purkinje dans une orientation rostrocaudal. La relation entre la cellule de Purkinje et le système de fibres à fibres moussues parallèle peut être caractérisée comme plusieurs-à-plusieurs. Avec la directivité étant orientation médio-latérale à l'intérieur de la couche moléculaire est à dire à angle droit par rapport aux dendrites des cellules de Purkinje, qui sont isoplanétique.
Le système de fibres à l'escalade
Originaire de l'olive inférieure controlatérale. À la suite de l'accouplement électrique entre les neurones olive inférieure, leur découplage dynamique via retour inhibition des noyaux cérébelleux et la topographie de la projection olivocerebellar, ce système génère synchrone (sur une échelle de temps en millisecondes) d'activation de pointe complexe de cellules de Purkinje, dans rostrocaudally bandes orientées. Ces bandes d'activité sont environ 250 m de large dans la direction médio-latérale, mais peut être de plusieurs millimètres de long dans le sens rostrocaudal et d'étendre les murs de la folia cérébelleux et à travers plusieurs lobules. La répartition du contrôle moteur instant en instant synchronie est dynamiquement modulée par l'olive inférieure avec le rôle majeur des afférences l'olive étant de déterminer le modèle de couplage électronique "effective" entre les neurones de l'olive et de ce fait la distribution de l'activité de pointe complexe synchrone travers le cortex cérébelleux. Changements dans les habitudes de synchronie sont associés à des mouvements effectués par les animaux effectuant une tâche motrice .. En effet. Le système de olivocerebellar peut être considéré comme un substrat malléable électriquement à partir de laquelle les synergies automobiles uniques peuvent être sculptées.
Le système de fibres à fibres moussues parallèle
Contrairement à la nature ponctuée d'activation du cervelet par le système olivocerebellar, le système de fibres à fibres parallèles mousse fournit une régulation continue et très délicate de l'excitabilité des noyaux cérébelleux , provoquée par l'activation de la tonique de pointes simples dans les cellules de Purkinje, qui génère finalement le contrôle fin du mouvement connu comme la coordination motrice. Le fait que les fibres moussues informer le cortex cérébelleux à la fois ascendante et descendante messages depuis et vers les centres moteurs de la moelle épinière et le tronc cérébral nous donne une idée du rôle ultime du système de fibres moussues: elle informe le cortex de la place et taux de mouvement des membres et met les intentions motrices générées par le cerveau dans le cadre de l'état de la masse au moment où le mouvement doit être exécuté. En outre, par ses effets sur les cellules inhibitrices GABAergiques cérébelleux nucléaires, qui projet à l'olive inférieure, il contribue à façonner le modèle de couplage entre les cellules de l'olive et donc la distribution de synchronie dans la prochaine décharge olivocerebellar.
Les noyaux cérébelleux
Les cellules de Purkinje sont la seule sortie du cortex cérébelleux et sont de nature inhibitrice Leurs axones en contact avec le cervelet et Deiters noyau vestibulaire comme leur seule cible. L'activité des noyaux cérébelleux est réglementée de trois façons: (1) par l'entrée excitateur de collatéraux des systèmes afférents cérébelleux, (2) par des entrées inhibitrices de cellules de Purkinje activés sur les voies de fibre de mousse, et (3) par des entrées de Purkinje les cellules activées par le système de fibres à l'escalade
Fonction cérébelleuse ensemble
La sortie du cervelet (les axones des noyaux cérébelleux) procéder pour générer l'activité de fond qui sert à définir le ton général et la posture qui donne le cortex moteur la capacité d'exécuter des mouvements sur la base de l'intention (la stratégie de mouvement). Dans ce contexte, le cervelet fournit les tactiques de l'activation musculaire multiples pour supporter ce type définissent mouvements. Et, tandis que le cerveau du moteur détermine où aller (impératif exécutif) le cervelet en œuvre son timing et module la force donnée à chaque commande de moteur, comme la coordination des mouvements est une fonction non-continue.
Dysfonctionnement
L'ataxie est un complexe de symptômes, impliquant généralement un manque de coordination, qui se trouve souvent dans les processus de maladies affectant le cervelet. Pour identifier les problèmes cérébelleux, les examen neurologique comprend l'évaluation de la marche (une démarche large étant indicatif de l'ataxie), des tests de pointer du doigt et de l'évaluation de la posture. Des anomalies structurales du cervelet (hémorragie, infarctus, cancer, la dégénérescence) peuvent être identifiés sur l'imagerie en coupe. Imagerie par résonance magnétique est la modalité de choix, que tomographie par ordinateur ne est pas suffisamment sensible pour détecter des anomalies structurales du cervelet.
Vieillissement
Un stéréologique étude a constaté que cérébelleuse humaine substance blanche a été réduit de 26% avec l'âge (plus de la tranche d'âge de 19 à 84). Les chercheurs de l'étude a pu détecter aucune perte globale de Purkinje ou cellules granulaires, mais dans le lobe antérieur il y avait une perte importante de ces types de cellules ainsi que d'une perte de volume de 30%. Avec imagerie par résonance magnétique une réduction volumétrique modérée avec l'âge vermis et l'hémisphère cérébelleux a été observée.
Une étude de autoradiographie du cervelet humain a trouvé une augmentation la liaison du H-3- KETANSERIN avec l'âge. (Kétansérine se lie principalement à la 5-HT 2A neurorécepteur) La même équipe de recherche n'a pas trouvé de corrélation significative avec l'âge dans leur d'homogénéiser étude de liaison. Un peu en ligne avec l'étude d'une autoradiographie étude de tomographie par émission de positons avec le altanserin récepteur de 5-HT 2A radioligand trouvé une corrélation positive entre l'âge et cérébelleux liaison non spécifique.
Les théories sur la fonction cérébelleuse
Deux théories principales portent sur la fonction du cervelet, à la fois face à la coordination motrice. On prétend que les fonctions du cervelet comme un régulateur de la "synchronisation des mouvements". Cela a émergé d'études de patients dont les mouvements sont perturbés chronométré.
La seconde, "Network Theory Tensor" fournit un modèle mathématique de la transformation des sensorielle (covariante) coordonne l'espace-temps dans le moteur (contravariant) coordonne par réseaux neuronaux cérébelleux.
Comme beaucoup de controverses dans les sciences physiques, il existe des preuves soutenant chacune des hypothèses ci-dessus. Les études de l'apprentissage moteur dans le réflexe vestibulo-oculaire et conditionné eyeblink démontrer que le calendrier et amplitude de mouvements tirés sont codées par le cervelet. Beaucoup mécanismes de plasticité synaptique ont été trouvés à travers le cervelet. Le modèle Marr-Albus attribue principalement l'apprentissage moteur à un mécanisme de plasticité unique: la dépression à long terme des synapses de fibres parallèles. La théorie Tensor Réseau des transformations sensori-motrices par le cervelet a également été expérimentalement soutenu.
Avec l'avènement de plus sophistiqué des techniques telles que la neuro-imagerie tomographie par émission de positons (PET), et de l'IRMf, de nombreuses fonctions diverses sont maintenant au moins partiellement attribués au cervelet. Ce qui était autrefois considéré comme une région principalement de l'intégration moteur / sensorielle se avère désormais être impliqués dans de nombreuses fonctions cognitives diverses.
La modélisation cérébelleuse
Comme mentionné dans la section précédente, il ya eu de nombreuses tentatives pour modéliser la fonction cérébelleuse. Les connaissances fournies par les modèles ont également conduit à des extrapolations dans les domaines de méthodes d'intelligence artificielle, en particulier les réseaux de neurones. Parmi les réalisations notables ont été Cerebellatron, cérébelleuse modèle associatif mémoire ou réseaux CCMC, et Spike FORCE pour le contrôle de mouvement robotique, et "Network Theory Tensor".
Images supplémentaires
- CT du cerveau de Mikael Häggström S3 I8.JPG
Tomodensitométrie de la tête, avec le cervelet visible à la partie inférieure.
Lobes

Diencéphale
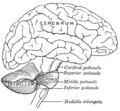
Schéma montrant les connexions de plusieurs parties du cerveau.

La surface supérieure du cervelet.

Sous surface du cervelet.

Coupe sagittale du cervelet, près de la jonction du vermis avec l'hémisphère.

Dissection montrant les fibres de projection du cervelet.

Schéma du toit du quatrième ventricule. La flèche est dans le trou de Magendie.

Dissection montrant le déroulement des fibres céphalo-rachidien.

Schéma montrant les positions des trois principaux citernes méningée.
Cervelet humain vue antérieure
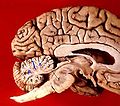
Cerveau humain vue médiosagittal