
Evolução
Sobre este escolas selecção Wikipedia
Esta seleção Escolas foi originalmente escolhido pelo SOS Children para as escolas no mundo em desenvolvimento sem acesso à internet. Ele está disponível como um download intranet. Com SOS Children você pode escolher para patrocinar crianças em mais de cem países
A evolução é a mudança na herdado características de biológico populações mais sucessiva gerações. Processos evolutivos dar origem a diversidade em todas as nível de organização biológica, incluindo espécies , organismos individuais e tais como moléculas de ADN e proteínas .
A vida na Terra evoluiu de uma ancestral comum universal aproximadamente 3,8 bilhões de anos atrás. Repetido especiação ea divergência de vida pode ser inferidas a partir de conjuntos comuns de características bioquímicas e morfológicas, ou compartilhada As sequências de ADN. Estes traços de homólogos e sequências são mais semelhantes entre as espécies que partilham um ancestral comum mais recente, e pode ser utilizado para reconstruir histórias evolutivas, usando ambas as espécies existentes eo registro fóssil . Padrões existentes de biodiversidade foram moldadas tanto por especiação e por extinção .
Charles Darwin e Alfred Wallace foram os primeiros a formular um argumento científico para a teoria da evolução por meio da seleção natural . Evolução por seleção natural é um processo que é inferida a partir de três fatos sobre populações: 1) mais descendentes são produzidos do que podem sobreviver, 2) traços variar entre os indivíduos, levando a diferentes taxas de sobrevivência e reprodução, e 3) as diferenças de característica são hereditárias. Assim, quando membros de uma população morrem, são substituídas pelo progênie de pais que foram melhor adaptado para sobreviver e se reproduzir no ambiente no qual a seleção natural ocorreu. Este processo cria e conserva características que são aparentemente equipada para o papéis funcionais que desempenham. A seleção natural é a única causa conhecida de adaptação, mas não a única causa conhecida de evolução. Outras causas, não adaptativas de evolução incluem e mutação deriva genética.
No início do século 20, genética foi integrado com a teoria da evolução pela seleção natural de Darwin através da disciplina de genética de populações. A importância da seleção natural como uma das causas da evolução foi aceito em outros ramos da biologia . Noções Além disso, anteriormente detidas cerca de evolução, tais como ortogênese e "Progresso" tornou- obsoleta. Os cientistas continuam a estudar vários aspectos da evolução através da formação e testar hipóteses, construindo teorias científicas, usando dados de observação e execução de em experiências tanto o campo e no laboratório. Biólogos concorda que a descendência com modificação é um dos mais estabelecida de forma confiável fatos na ciência. Descobertas em biologia evolutiva fez um impacto significativo não apenas dentro dos ramos tradicionais da biologia, mas também em outras disciplinas acadêmicas (por exemplo, antropologia e psicologia) e na sociedade em geral.
História do pensamento evolutivo
A proposta de que um tipo de animal pode descer de um animal de um outro tipo de volta a algum do primeiro filósofos gregos pré-socráticos, como Anaximandro e Empédocles. Em contraste com estes visões materialistas, Aristóteles compreendeu todas as coisas naturais, não só vivem as coisas, como ser imperfeito actualisations de diferentes possibilidades naturais fixos, conhecidos como " formas "," idéias ", ou (em latim traduções)" espécie ". Esta era parte de sua compreensão teleológica natureza em que todas as coisas têm um papel a intenção de jogar em um divino ordem cósmica. O romano poeta e filósofo Tito Lucrécio Carus propôs a possibilidade de mudanças evolucionárias de organismos. As variações desta ideia tornou-se a compreensão padrão da Idade Média , e foram integrados de aprendizagem cristã, mas Aristóteles não exigir que os tipos reais de animais correspondeu um-para-um com formas metafísicas exatas e, especificamente, deu exemplos de como novos tipos de seres vivos poderia vir a ser.
No século 17 o novo método de ciência moderna rejeitou a abordagem de Aristóteles, e procurou explicações de fenômenos naturais em termos de leis da natureza que eram as mesmas para todas as coisas visíveis, e não precisam de assumir quaisquer categorias naturais fixos, nem qualquer ordem cósmica divina. Mas esta nova abordagem demorou a se enraizar nas ciências biológicas, que se tornaram o último bastião do conceito de tipos naturais fixos. John Ray utilizado um dos termos mais gerais anteriormente para tipos naturais fixos, "espécies", a aplicar aos tipos de animais e plantas, mas ao contrário de Aristóteles ele identificou estritamente cada tipo de coisa viva, como espécie, e propôs que cada espécie pode ser definida pelas características que se perpetuam cada geração. Estas espécies foram designados por Deus, mas que mostram diferenças causadas por condições locais. A classificação biológica introduzidas por Carolus Linnaeus em 1735, também viram espécies como fixado de acordo com um plano divino.

Outros naturalistas deste tempo especulou sobre a mudança evolutiva das espécies ao longo do tempo de acordo com as leis naturais. Maupertuis escreveu em 1751 de modificações naturais que ocorrem durante a reprodução e acumulando ao longo de muitas gerações para produzir novas espécies. Buffon sugeriu que as espécies poderiam degenerar em diferentes organismos, e Erasmus Darwin propôs que todos os animais de sangue quente poderia ter descendem de um único micro-organismo (ou "filamento"). O primeiro esquema evolutivo de pleno direito foi A teoria de Lamarck "transmutação" de 1809 que previa geração espontânea produzindo continuamente formas de vida simples desenvolvido maior complexidade em linhagens paralelas com uma tendência progressiva inerente, e que essas linhagens em um nível local adaptadas ao ambiente por mudanças herdando causados pelo uso ou desuso pais. (O último processo foi mais tarde chamado lamarckismo.) Essas idéias foram condenados pelos naturalistas estabelecidos como a especulação falta de apoio empírico. Em particular Georges Cuvier insistiu que as espécies não foram relacionados e fixa, suas semelhanças refletindo desígnio divino para as necessidades funcionais. Nesse meio tempo, as idéias de Ray do projeto benevolente tinha sido desenvolvido por William Paley numa teologia natural que propôs adaptações complexas como evidência de desígnio divino, e era admirado por Charles Darwin.
A ruptura crítica do conceito de espécies fixas em biologia começou com a teoria da evolução pela seleção natural, que foi formulada por Charles Darwin. Parcialmente influenciado pela Um Ensaio sobre o Princípio da População por Thomas Robert Malthus , Darwin observou que o crescimento da população levaria a uma "luta pela existência" onde as variações favoráveis poderia prevalecer como outros pereceram. Cada geração, muitos filhos não conseguem sobreviver a uma idade de reprodução por causa de recursos limitados. Isso poderia explicar a diversidade de animais e plantas a partir de um ancestral comum por meio do trabalho das leis naturais que trabalham o mesmo para todos os tipos de coisa. Darwin estava desenvolvendo sua teoria da " seleção natural "de 1838 em diante, até Alfred Russel Wallace lhe enviou uma teoria semelhante em 1858. Ambos os homens apresentaram o seu documentos separados para o Linnean Society of London. No final de 1859, publicação de Darwin A Origem das Espécies explicou seleção natural em detalhe e de uma forma que levou a uma cada vez mais ampla aceitação do A evolução darwiniana. Thomas Henry Huxley aplicado idéias de Darwin para os seres humanos, usando paleontologia e anatomia comparativa para fornecer fortes evidências de que os seres humanos e macacos compartilhado um ancestral comum. Alguns foram perturbado por este, uma vez que implica que os seres humanos não têm um lugar especial no universo.
Mecanismos precisos de herdabilidade reprodutiva e a origem de novos traços permanece um mistério. Para este fim, Darwin desenvolveu sua teoria provisória do pangênese. Em 1865 Gregor Mendel informou que as características eram herdadas de uma maneira previsível através da variedade independente e segregação de elementos (mais tarde conhecido como genes). As leis de Mendel da herança acabou por suplantar a maioria de teoria pangênese de Darwin. August Weismann fez a distinção importante entre células germinativas (espermatozóides e óvulos) e células somáticas do corpo, o que demonstra que a hereditariedade passa somente através da linha germinal. Hugo de Vries conectado teoria de Darwin para pangênese germe / distinção célula soma de Weismann e propôs que pangenes de Darwin foram concentrados no núcleo da célula e quando expressa que poderiam passar para o citoplasma para alterar a estrutura das células. De Vries também foi um dos investigadores que fizeram o trabalho de Mendel bem conhecida, acreditando que as características mendelianas correspondeu à transferência de variações hereditárias ao longo da linha germinal. Para explicar como as novas variantes são originários, De Vries desenvolveu um teoria da mutação que levou a uma ruptura temporária entre aqueles que aceitaram a evolução darwiniana e biometristas que aliados de Vries. Na virada do século 20, pioneiros no campo da genética de populações, tais como JBS Haldane, Sewall Wright, e Ronald Fisher, definir as bases da evolução para uma filosofia estatística robusta. A falsa contradição entre a teoria de Darwin, mutações genéticas, e herança mendeliana foi, assim, reconciliado.
Na década de 1920 e 1930 um síntese evolutiva moderna conectado seleção natural, teoria da mutação, e herança mendeliana em uma teoria unificada que aplicado geralmente a qualquer ramo da biologia. A síntese moderna foi capaz de explicar os padrões observados em várias espécies em populações, através de transições fósseis em paleontologia e mecanismos celulares até mesmo em complexas biologia do desenvolvimento. A publicação da estrutura do DNA por James Watson e Francis Crick em 1953 demonstrou uma base física para a herança. A biologia molecular melhorado nossa compreensão da relação entre genótipo e fenótipo. Os avanços também foram feitas em filogenética sistemática, mapeando a transição de traços em um quadro comparativo e testável através da publicação e da utilização de árvores evolucionárias. Em 1973, o biólogo evolucionista Theodosius Dobzhansky escreveu que "nada em biologia faz sentido exceto à luz da evolução", porque ele trouxe à luz as relações de que primeiro parecia fatos desconexos da história natural em um coerente explicativo corpo de conhecimento que descreve e prediz muitos fatos observáveis sobre a vida neste planeta.
Desde então, a síntese moderna foi prorrogado para explicar fenômenos biológicos em toda a escala completa e integradora da hierarquia biológica, a partir de genes de espécies. Esta extensão foi apelidado de " eco-evo-devo ".
Hereditariedade

Evolução em organismos ocorre através de mudanças na hereditárias traços - características particulares de um organismo. Nos seres humanos, por exemplo, cor dos olhos é uma característica herdada e um indivíduo pode herdar o "traço de olhos marrom" de um de seus pais. Herdados características são controladas pelos genes e o conjunto completo de genes dentro de um organismo de genoma é chamado de genótipo.
O conjunto completo de características observáveis que compõem a estrutura eo comportamento de um organismo é chamado de fenótipo. Essas características da interacção do seu genótipo com o ambiente. Como resultado, muitos aspectos do fenótipo de um organismo não são herdadas. Por exemplo, pele bronzeada vem da interação entre genótipo e da luz solar de uma pessoa; assim, suntans não são repassados para as crianças das pessoas. No entanto, algumas pessoas se bronzear mais facilmente do que outros, devido a diferenças no seu genótipo; um exemplo notável são pessoas com o traço herdado de albinismo , que não fazem tan em tudo e são muito sensíveis à queimaduras solares.
Características hereditárias são transmitidas de uma geração para a outra via DNA , uma molécula que codifica a informação genética. ADN é um longo polímero composto por quatro tipos de bases. A sequência de bases ao longo de uma molécula de DNA particular especificar a informação genética, de um modo semelhante a uma sequência de letras soletrar uma frase. Antes de uma célula se divide, o DNA é copiado, de modo que cada uma das duas células resultantes irá herdar a sequência de ADN. As porções de uma molécula de DNA que especificam uma única unidade funcional são chamados genes; diferentes genes têm diferentes sequências de bases. Dentro de células , os longos fios de forma condensada estruturas de DNA chamado cromossomas. O local específico de uma sequência de ADN dentro de um cromossoma é conhecido como um lócus. Se a sequência de ADN num locus varia entre os indivíduos, as diferentes formas de esta sequência são chamados alelos. Sequências de DNA podem mudar através mutações, produzindo novos alelos. Se uma mutação ocorre dentro de um gene, o novo alelo pode afectar a característica de que os controlos do gene, que altera o fenótipo do organismo. No entanto, enquanto este simples correspondência entre um alelo e uma característica funciona em alguns casos, a maioria das características são mais complexos e são controlados pela vários genes que interagem.
Descobertas recentes têm confirmado exemplos importantes de mudanças hereditárias que não podem ser explicadas por alterações à sequência de nucleotídeos no DNA. Estes fenómenos são classificados como sistemas de herança epigenética. DNA marcação metilação cromatina, laços metabólicas auto-sustentável, o silenciamento de genes por RNA de interferência e os três dimensional conformação das proteínas (como prions ) são áreas onde os sistemas de herança epigenética foram descobertos em nível organísmico. Biólogos do desenvolvimento sugerem que as interações complexas em redes genéticas e comunicação entre as células pode levar a variações hereditárias que podem subjacentes alguns dos mecânica em e plasticidade de desenvolvimento canalização. Hereditariedade também pode ocorrer em escalas ainda maiores. Por exemplo, a herança ecológica através do processo de nicho de construção é definido pelas atividades regulares e repetidos de organismos no seu meio ambiente. Isso gera um legado de efeitos que modificam e se alimentam de volta para o regime de selecção das gerações subsequentes. Descendentes herdam genes além de características ambientais gerados pelas ações ecológicas dos antepassados. Outros exemplos de herdabilidade em evolução que não estão sob o controle direto de genes incluem a herança de traços culturais e simbiogênese.
Variação


De um organismo individual resultados de fenótipo tanto a sua genótipo e a influência do ambiente em que vive. Uma parte substancial da variação de fenótipos em uma população é causada pelas diferenças entre seus genótipos. O síntese evolutiva moderna define evolução como a mudança ao longo do tempo nesta variação genética. A frequência de um alelo particular irá tornar-se mais ou menos prevalente em relação a outras formas do mesmo gene. Variação desaparece quando um novo alelo atinge o ponto de fixação - quando se quer desaparece da população ou substitui o alelo ancestral inteiramente.
A seleção natural só vai provocar o desenvolvimento se não houver o suficiente variação genética numa população. Antes da descoberta de Genética mendeliana, uma hipótese comum foi herança de mistura. Mas com mistura de herança, variância genética seria rapidamente perdido, tornando evolução por seleção natural implausível. O Princípio de Hardy-Weinberg fornece a solução de como a variação é mantida numa população com Herança mendeliana. As frequências de alelos (as variações de um gene) permanecerá constante na ausência de selecção, mutação, a migração e a deriva genética.
Variação vem mutações em material genético, através de rearranjo de genes a reprodução sexual e a migração entre as populações ( o fluxo de genes). Apesar da introdução de novo constante variação através da mutação do gene e o fluxo, a maior parte do genoma de uma espécie é idêntico em todos os indivíduos dessa espécie. No entanto, mesmo relativamente pequenas diferenças no genótipo pode conduzir a diferenças dramáticas no fenótipo: por exemplo, chimpanzés e seres humanos diferem em apenas cerca de 5% dos seus genomas.
Mutação

As mutações são alterações na sequência de ADN do genoma de uma célula. Quando ocorrem mutações, eles podem não ter efeito, alterar a produto de um gene, ou impedir o funcionamento do gene. Com base em estudos da mosca Drosophila melanogaster , foi sugerido que, se uma mutação muda de uma proteína produzida por um gene, isto irá provavelmente ser prejudicial, com cerca de 70% destas mutações que têm efeitos prejudiciais, e sendo o restante neutro ou fracamente benéfica.
As mutações podem envolver grandes seções de um cromossomo a tornar-se duplicado (geralmente por recombinação genética), que pode introduzir cópias adicionais de um gene num genoma. Cópias extras de genes são uma importante fonte de matéria-prima necessária para novos genes para evoluir. Isto é importante porque a maioria dos novos genes evoluir dentro famílias de genes de genes pré-existentes que compartilham ancestrais comuns. Por exemplo, o olho humano utiliza quatro genes para fazer estruturas que detectam luz: três para visão de cores e uma para visão noturna; todos os quatro são descendentes de um único gene ancestral.
Novos genes pode ser gerado a partir de um gene ancestral, quando uma cópia sofre mutação duplicado e adquire uma nova função. Este processo é fácil uma vez que um gene tiver sido duplicada, porque aumenta a redundância do sistema; um gene do par pode adquirir uma nova função enquanto a outra cópia continua a desempenhar a sua função original. Outros tipos de mutações podem até mesmo gerar inteiramente novos genes de DNA não-codificante anteriormente.
A geração de novos genes também pode envolver as peças pequenas de vários genes a ser duplicada, com estes fragmentos, em seguida, a recombinação para formar novas combinações com novas funções. Quando os novos genes são montados a partir de partes pré-baralhar existentes, domínios actuar como módulos com funções independentes simples, que podem ser misturados em conjunto para produzir novas combinações com funções novas e complexas. Por exemplo, sintases de policétido são grandes enzimas que produzem antibióticos; eles contêm até cem domínios independentes que catalisam cada uma das etapas do processo global, como um passo na linha de montagem.
Sexo e recombinação
Em organismos de reprodução assexuada, os genes são herdados juntos, ou ligados, pois eles não podem se misturar com genes de outros organismos durante a reprodução. Em contraste, a descendência de organismos sexuais contêm misturas aleatórias de cromossomos dos pais que são produzidos através de sortimento independente. Em um processo relacionado chamado recombinação homóloga, organismos sexuais trocar DNA entre dois cromossomos correspondentes. A recombinação e o rearranjo não alteram as frequências alélicas, mas em vez disso mudar alelos que são associados uns com os outros, a produção de descendência com novas combinações de alelos. Sexo geralmente aumenta a variação genética e podem aumentar a taxa de evolução.
O fluxo gênico
O fluxo gênico é a troca de genes entre populações e entre espécies. Pode, portanto, ser uma fonte de variação que é novo para uma população ou para uma espécie. O fluxo de genes pode ser causado pelo movimento de indivíduos entre populações separadas de organismos, como pode ser causado pelo movimento dos ratos entre o interior e populações costeiras, ou o movimento de pólen entre tolerante heavy metal e heavy metal populações sensíveis de gramíneas.
A transferência de genes entre espécies inclui a formação de organismos híbridos e transferência de genes horizontal. Transferência de genes horizontal é a transferência de material genético de um organismo para outro organismo que não é a sua prole; este é o mais comum entre bactérias . Na medicina, isso contribui para a propagação de resistência aos antibióticos, como quando uma bactéria adquire genes de resistência que pode transferi-las rapidamente para outras espécies. Transferência horizontal de genes a partir de bactérias para eucariotas tais como leveduras Saccharomyces cerevisiae eo feijão azuki besouro Callosobruchus chinensis ocorreu. Um exemplo das transferências de maior escala são a eucariótica rotíferos bdelloid, que receberam uma série de genes de bactérias, fungos e plantas. Os vírus também podem transportar DNA entre organismos, permitindo a transferência de genes mesmo entre domínios biológicos.
Transferência de genes em larga escala também ocorreu entre os ancestrais de células eucarióticas e bactérias , durante a aquisição de cloroplastos e mitocôndrias . É possível que os próprios eucariotas originado a partir de transferências de genes horizontais entre as bactérias e archaea.
Mecanismos
De um Perspectiva neo-darwinista, a evolução ocorre quando há mudanças nas frequências de alelos dentro de uma população de cruzamentos organismos. Por exemplo, o alelo de cor preta numa população de traças cada vez mais comum. Mecanismos que podem levar a alterações nas freqüências alélicas incluem a seleção natural , deriva genética, hitchhiking genética, e mutação fluxo gênico.
Seleção natural
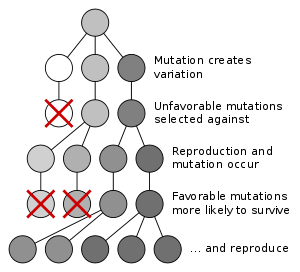
Evolução por meio de seleção natural é o processo pelo qual as mutações genéticas que melhoram a reprodução tornam-se e permanecem mais comuns em gerações sucessivas de uma população. Tem sido frequentemente chamado um mecanismo de "auto-evidente" porque segue necessariamente a partir de três simples fatos:
- Variação hereditária existe dentro de populações de organismos.
- Organismos produzem mais descendência do que pode sobreviver.
- Estes filhotes variam em sua capacidade de sobreviver e se reproduzir.
Estas condições produzem a concorrência entre os organismos para a sobrevivência e reprodução. Consequentemente, os organismos com características que lhes dão uma vantagem sobre seus concorrentes passar essas características vantajosas no, enquanto traços que não conferem uma vantagem que não são passados para a próxima geração.
O conceito central da seleção natural é o aptidão evolutiva de um organismo. Educação Física é medido pela capacidade de um organismo para sobreviver e se reproduzir, o que determina o tamanho da sua contribuição genética para a próxima geração. No entanto, a aptidão não é o mesmo que o número total de prole: em vez de fitness é indicado pela proporção de gerações subsequentes que são portadores de genes de um organismo. Por exemplo, se um organismo poderia sobreviver bem e reproduzem-se rapidamente, mas sua prole foram todos muito pequeno e fraco para sobreviver, este organismo faria pouca contribuição genética para as gerações futuras e teria, assim, baixa aptidão.
Se um alelo aumenta a aptidão mais do que os outros alelos do mesmo gene, em seguida, a cada geração esse alelo vai se tornar mais comum no seio da população. Estes traços são disse a ser "selecionado para". Exemplos de características que podem aumentar a aptidão são reforçadas sobrevivência e aumentou fecundidade. Por outro lado, a menor aptidão causado por ter um alelo menos benéficos ou deletérios resultado nesta alelo se tornando mais raro - eles são "selecionadas contra". É importante ressaltar que a adequação de um alelo não é uma característica fixa; se o ambiente muda, anteriormente traços neutros ou nocivos podem tornar-se características benéficas e previamente benéficos tornam-se prejudiciais. No entanto, mesmo que a direção da seleção faz reverter desta forma, características que foram perdidas no passado, não pode voltar a evoluir de forma idêntica (ver A lei de Dollo).

A selecção natural dentro de uma população de uma característica que pode variar dentro de um intervalo de valores, tais como a altura, podem ser classificados em três tipos diferentes. O primeiro é selecção direccional, que é uma mudança no valor médio de uma característica ao longo do tempo - por exemplo, organismos lentamente a ficar mais alto. Em segundo lugar, seleção disruptiva é seleção para valores extremos das características e muitas vezes resulta em dois valores diferentes se tornar mais comum, com selecção em relação ao valor médio. Isto seria quando os organismos ou baixos ou altos tinham uma vantagem, mas não os de estatura mediana. Finalmente, em estabilização selecção existe selecção contra valores extremos das características em ambas as extremidades, o que provoca uma diminuição na variação em torno do valor médio e menos diversidade. Este seria, por exemplo, fazer com que os organismos para se tornar lentamente todos a mesma altura.
Um caso especial da seleção natural é seleção sexual, que é a seleção de qualquer característica que aumenta sucesso de acasalamento, aumentando a atratividade de um organismo para potenciais companheiros. Traços que evoluíram através da seleção sexual são particularmente proeminentes nos machos de algumas espécies de animais, apesar de características como chifres pesados, chamados de acasalamento ou cores brilhantes que atraem predadores, diminuindo a sobrevivência de machos individuais. Esta desvantagem sobrevivência é equilibrada pela maior sucesso reprodutivo em homens que mostram estes difícil de falsificar, sexualmente traços selecionado.
A seleção natural mais geralmente faz com que a natureza a medida contra a qual os indivíduos e as características individuais, são mais ou menos chances de sobreviver. "Nature", nesse sentido, refere-se a um ecossistema, isto é, um sistema no qual os organismos interagem com todos os outros elementos, física, bem como biológico, na sua locais ambiente. Eugene Odum, um dos fundadores da ecologia, definiu um ecossistema como: "Qualquer unidade que inclui todos os organismos ... em uma determinada área interagindo com o ambiente físico, de modo que um fluxo de energia leva a claramente definidos estrutura trófica, a diversidade biótica e ciclos de materiais (ou seja: troca de materiais entre vivos e não vivos peças) dentro do sistema. " Cada população dentro de um ecossistema ocupa uma distinta nicho, ou posição, com relações distintas para outras partes do sistema. Esses relacionamentos envolvem a história de vida do organismo, a sua posição no cadeia alimentar e sua distribuição geográfica. Este amplo entendimento da natureza permite aos cientistas traçar as forças específicas que, juntos, compõem a seleção natural.
A seleção natural pode atuar em diferentes níveis de organização, tais como genes, células, organismos individuais, grupos de organismos e espécies. A seleção pode agir em vários níveis simultaneamente. Um exemplo de selecção que ocorre abaixo do nível do organismo individual são chamados genes transposons, que podem se replicar e se espalham ao longo de um genoma. Selecção a um nível acima do indivíduo, tal como selecção grupo, pode permitir a evolução de cooperação, tal como discutido abaixo.
Mutação tendenciosa
Para além de ser uma das principais fontes de variação, a mutação pode também funcionar como um mecanismo de evolução quando existem diferentes probabilidades a nível molecular para diferentes mutações de ocorrer, um processo conhecido como viés mutação. Se dois genótipos, por exemplo um com o nucleótido G e outro com o nucleótido A na mesma posição, a mesma tem de fitness, mas a mutação de G para A acontece mais frequentemente do que a mutação de A para G, em seguida, com genótipos Um tenderá a evoluir. Inserção diferente vs. preconceitos apagamento de mutação em diferentes taxa pode levar à evolução de diferentes tamanhos de genoma. Desvios do desenvolvimento ou mutacionais também têm sido observadas em evolução morfológica. Por exemplo, de acordo com o fenótipo-primeira teoria da evolução, as mutações podem, eventualmente, causar a assimilação genética de características que antes eram induzida pelo ambiente.
Efeitos de polarização mutação estão sobrepostos a outros processos. Se selecção favoreceria uma ou de duas mutações, mas não há nenhuma vantagem adicional de que tem ambos, em seguida, a mutação que ocorre com maior frequência é o de que é mais provável de se fixar numa população. As mutações que conduzem à perda da função de um gene são muito mais comuns do que as mutações que produzem um gene novo, completamente funcional. Mais mutações de perda de função são selecionadas contra. Mas quando a seleção é fraca, o viés de mutação para a perda de função podem afetar a evolução. Por exemplo, os pigmentos não são mais úteis quando os animais vivem na escuridão da caverna, e tendem a ser perdidos. Este tipo de perda de função possam ocorrer devido a polarização mutação, e / ou porque a função tinha um custo, e uma vez que o benefício da função desapareceu, a selecção natural leva à perda. Perda de capacidade de esporulação numa bactéria durante a evolução laboratório parece ter sido causada por mutação viés, em vez de a seleção natural contra o custo de manter a capacidade de esporulação. Quando não há nenhuma selecção para a perda de função, a velocidade a que se desenvolve a perda depende mais da taxa de mutação do que na tamanho efetivo da população, indicando que ele é mais influenciado pelo viés mutação que pela deriva genética.
A deriva genética

A deriva genética é a mudança na freqüência do alelo de uma geração para a seguinte, que ocorre porque os alelos estão sujeitos a erro de amostragem. Como resultado, quando forças seletivas estão ausentes ou relativamente fraca, as freqüências alélicas tendem a "deriva" cima ou para baixo de forma aleatória (em um passeio aleatório). Essa deriva pára quando um alelo eventualmente torna-se fixo, quer por desaparecer da população, ou substituir os outros alelos inteiramente. A deriva genética pode, portanto, eliminar alguns alelos de uma população devido ao acaso. Mesmo na ausência de forças seletivas, deriva genética pode causar duas populações separadas que começaram com a mesma estrutura genética a se afastar em duas populações divergentes com diferentes conjuntos de alelos.
Geralmente é difícil medir a importância relativa dos processos de seleção e neutras, incluindo deriva. A importância comparativa das forças adaptativos e não adaptativos como motores da mudança evolutiva é uma área de pesquisa atual .
O teoria neutra da evolução molecular proposto que a maioria das mudanças evolutivas são o resultado da fixação de mutações neutras por deriva genética. Assim, neste modelo, a maioria das alterações genéticas em uma população são o resultado de mutação pressão constante e deriva genética. Esta forma da teoria neutra é agora amplamente abandonada, uma vez que não parece enquadrar-se a variação genética visto na natureza. No entanto, uma versão mais recente e mais bem apoiada deste modelo é o teoria quase neutro, em que uma mutação que seria neutro numa pequena população não é necessariamente neutro numa grande população. Outras teorias alternativas propõem que deriva genética é ofuscado por outras forças estocásticos em evolução, como hitchhiking genética, também conhecida como projecto de genética.
O tempo para um alelo neutro para se fixar por deriva genética depende do tamanho da população, com a fixação ocorre mais rapidamente em populações menores. O número de indivíduos de uma população não é crítica, mas em vez disso uma medida conhecida como o tamanho efetivo da população. A população eficaz é geralmente menor do que a população total, uma vez que leva em conta factores tais como o nível de endogamia e a fase do ciclo de vida em que a população é a menor. O tamanho eficaz da população podem não ser os mesmos para todos os genes na mesma população.
Hitchhiking genética
A recombinação permite alelos na mesma fita de DNA para se separar. No entanto, a taxa de recombinação é baixa (cerca de dois eventos por cromossoma por geração). Como resultado, os genes juntas num cromossoma pode nem sempre ser baralhado de distância um do outro e os genes que estão próximos uns dos outros tendem a ser herdados em conjunto, um fenómeno conhecido como ligação. Esta tendência é medida encontrando a frequência com dois alelos ocorrem juntos em um único cromossomo em comparação com expectativas, que é chamado sua desequilíbrio de ligação. Um conjunto de alelos que é geralmente herdado de um grupo é um chamado haplótipo. Isso pode ser importante quando um alelo em um haplótipo particular é fortemente benéfico: a seleção natural pode conduzir um limpeza seletiva que também fará com que os outros alelos do haplótipo para tornar-se mais comuns na população; este efeito é chamado hitchhiking genética ou projecto de genética.Projecto genética causada pelo facto de alguns genes são neutros geneticamente ligado a outros que estão sob selecção pode ser parcialmente captada por uma população de tamanho eficaz apropriada.
O fluxo gênico
O fluxo gênico é a troca de genes entre populações e entre espécies. A presença ou ausência de fluxo gênico muda fundamentalmente o curso da evolução. Devido à complexidade dos organismos, quaisquer duas populações completamente isolado eventualmente evoluir incompatibilidades genéticos através de processos neutros, como no modelo de Bateson-Dobzhansky-Muller, mesmo se ambas as populações permanecem essencialmente idênticos em termos da sua adaptação ao ambiente.
Se a diferenciação genética entre as populações desenvolve, o fluxo gênico entre populações podem apresentar características ou alelos, que são desvantajosos na população local e isso pode levar a essas populações organismo dentro de evoluir mecanismos que impedem que acoplam-se com populações geneticamente distantes, acabou resultando no aparecimento de novo espécies. Assim, a troca de informação genética entre indivíduos é fundamentalmente importante para o desenvolvimento do conceito de espécie biológica (BSC).
Durante o desenvolvimento da síntese moderna, Sewall Wright desenvolveu sua teoria equilíbrio mudança que o fluxo gênico entre as populações parcialmente isoladas foi um aspecto importante da evolução adaptativa. No entanto, recentemente tem havido crítica substancial da importância da teoria mudança no equilíbrio.
Outcomes
Evolução influencia todos os aspectos da forma e do comportamento dos organismos. O mais proeminente são as comportamentais e físicas específicas adaptações que são o resultado da seleção natural. Essas adaptações aumentam a aptidão por auxiliando atividades como encontrar comida, evitar predadores ou atrair parceiros. Organismos também podem responder à seleção por cooperando uns com os outros, geralmente por ajudar os seus parentes ou se engajar em mutuamente benéfica simbiose. A mais longo prazo, a evolução produz novas espécies através populações ancestrais de divisão de organismos em novos grupos que não podem ou não vai cruzar.
Estes resultados de evolução são, por vezes, divididos em macro-evolução, o qual é a evolução que ocorre igual ou superior ao nível de espécies, tais como a extinção e especiação e microevolution, que é alterações evolutivas mais pequenas, tais como adaptações, dentro de uma espécie ou população. Em geral, macroevolução é considerada como o resultado de longos períodos de microevolution. Assim, a distinção entre micro e macroevolução não é essencial um - a diferença é simplesmente o tempo envolvido. No entanto, em macroevolution, as características de toda a espécie pode ser importante. Por exemplo, uma grande quantidade de variação entre indivíduos permite que uma espécie se adaptar rapidamente aos novos habitats, diminuindo a chance de se tornar extinta, enquanto uma distribuição geográfica ampla aumenta a chance de especiação, tornando-o mais provável que parte da população vai tornar-se isolado. Neste sentido, microevolução e macroevolução pode envolver a seleção em níveis diferentes - com microevolução agindo sobre genes e organismos, contra processos macroevolutivos tais como a seleção de espécies agindo em espécies inteiras e afetando suas taxas de especiação e extinção.
Um equívoco comum é que a evolução tem metas ou planos de longo prazo; realisticamente no entanto, a evolução não tem meta de longo prazo e não produz necessariamente uma maior complexidade. Embora espécies complexas evoluíram, eles ocorrer como um efeito colateral do número total de organismos e crescentes formas de vida simples ainda permanecem mais comum na biosfera. Por exemplo, a esmagadora maioria das espécies são microscópicas procariontes, que formam cerca de metade do mundo biomassa apesar de seu tamanho pequeno, e constituem a grande maioria da biodiversidade da Terra. Organismos simples têm sido, portanto, a forma dominante de vida na Terra ao longo de sua história e continuam a ser a principal forma de vida até os dias de hoje, com a vida complexo só aparecendo mais diversificado, porque é mais perceptível. Com efeito, a evolução do microorganismos é particularmente importante para a pesquisa evolutiva moderna , uma vez que a sua reprodução rápida permite o estudo da evolução experimental ea observação de evolução e adaptação em tempo real.
Adaptação

A adaptação é o processo que faz com que os organismos mais adequado para a sua habitat. Além disso, a adaptação termo pode referir-se a uma característica que é importante para a sobrevivência de um organismo. Por exemplo, a adaptação dos dentes dos cavalos para a moagem de grama. Ao usar o termo adaptação para o processo evolutivo e característica adaptativa para o produto (a parte do corpo ou função), os dois sentidos da palavra pode ser distinto. Adaptações são produzidas por seleção natural . As definições a seguir são devido a Theodosius Dobzhansky.
- Adaptaçãoé o processo evolutivo pelo qual um organismo se torna mais capaz de viver em seuhabitat ou habitats.
- Adaptabilidadeé o estado de ser adaptada: o grau em que um organismo pode viver e reproduzir-se em um dado conjunto de habitats.
- Umacaracterística de adaptaçãoé um aspecto do padrão de desenvolvimento do organismo que se permite ou aumenta a probabilidade de que o organismo sobreviver e reprodução.
A adaptação pode causar tanto o ganho de um novo recurso, ou a perda de uma característica ancestral. Um exemplo mostra que ambos os tipos de mudança é adaptação bacteriana a antibiótico de selecção, com alterações genéticas que causam a resistência aos antibióticos por tanto modificar o destino do fármaco, ou o aumento da actividade de transportadores que bombeiam o fármaco para fora da célula. Outros exemplos notáveis são os de bactérias de Escherichia coli em evolução da capacidade de utilizar o ácido cítrico como um nutriente de uma experiência de laboratório a longo prazo, Flavobacterium evoluir uma nova enzima que permite que estas bactérias a crescer nos subprodutos de fabrico de nylon, e a bactéria do solo Sphingobium evoluindo uma inteiramente nova via metabólica que degrada o sintético pesticida pentaclorofenol. Uma idéia interessante, mas ainda controverso é que algumas adaptações podem aumentar a capacidade de organismos para gerar diversidade genética e adaptar-se pela seleção natural (aumentando "organismos evolvability).

Adaptação ocorre através da modificação gradual das estruturas existentes. Consequentemente, as estruturas com organização interna semelhante podem ter diferentes funções em organismos relacionados. Este é o resultado de uma única estrutura ancestral ser adaptado para funcionar de diferentes maneiras. Os ossos dentro bastão asas, por exemplo, são muito semelhantes aos de ratos pés e as mãos de primatas, devido à descida de todas estas estruturas a partir de um antepassado comum de mamífero. No entanto, uma vez que todos os organismos vivos estão relacionadas, em certa medida, mesmo em órgãos que parecem ter pouca ou nenhuma semelhança estrutural, tais como artrópodes , lulas e olhos vertebrados, ou os membros e as asas de artrópodes e vertebrados, pode depender de um conjunto comum de genes homólogos que controlam a sua montagem e função; esta é chamada homologia profunda.
Durante a evolução, algumas estruturas podem perder sua função original e tornar-se estruturas vestigiais . Tais estruturas podem ter pouca ou nenhuma função em uma espécie atual, ainda tem uma função clara em espécies ancestrais, ou outras espécies estreitamente relacionadas. Exemplos incluem pseudogenes, os restos não-funcionais de olhos em peixe cego da caverna-moradia, asas em aves que não voam, e a presença de ossos do quadril em baleias e cobras. Exemplos de estruturas vestigiais em humanos incluem os dentes do siso, o cóccix, o apêndice vermiforme, e outros vestígios comportamentais, tais como arrepios e reflexos primitivos.
No entanto, muitos traços que parecem ser simples adaptações são de fato exaptações: estruturas originalmente adaptados para uma função, mas que coincidentemente se tornaram um tanto útil para alguma outra função no processo. Um exemplo é o lagarto Africano holaspis guentheri , que desenvolveu uma cabeça extremamente plana para se esconder em fendas, como pode ser visto olhando para seus parentes próximos. No entanto, nesta espécie, a cabeça se tornou tão achatado que auxilia no deslizamento de árvore em árvore-an exaptação. Dentro das células, máquinas moleculares, tais como o bacteriana e flagelos máquinas proteína triagem evoluído pelo recrutamento de várias proteínas pré-existente que anteriormente tinham diferentes funções. Outro exemplo é o recrutamento de enzimas de glicólise e metabolismo xenobiótico para servir como proteínas estruturais chamados cristalinas dentro das lentes dos organismos de olhos .
Um princípio importante da ecologia é a de exclusão competitiva: há duas espécies podem ocupar o mesmo nicho no mesmo ambiente por um longo tempo. Consequentemente, a seleção natural tenderá a forçar espécies de se adaptar a diferentes nichos ecológicos. Isso pode significar que, por exemplo, duas espécies de peixe ciclídeo adaptar-se a viver em diferentes habitats, que irá minimizar a concorrência entre eles para o alimento.
Uma área de investigação em curso em biologia evolutiva do desenvolvimento é a base de desenvolvimento de adaptações e exaptações. Esta pesquisa aborda a origem e evolução do desenvolvimento embrionário e como as modificações de desenvolvimento e processos de desenvolvimento produzem novas características. Estes estudos têm mostrado que a evolução pode alterar o desenvolvimento para produzir novas estruturas, tais como estruturas ósseas embrionárias que se desenvolvem na mandíbula em outros animais vez que fazem parte do ouvido médio em mamíferos. É também possível para as estruturas que foram perdidos na evolução para reaparecer devido a alterações nos genes de desenvolvimento, como uma mutação em galinhas causando embriões para crescer dentes semelhantes aos de crocodilos . Ele agora está se tornando claro que a maioria das alterações na forma de organismos são devido a alterações em um pequeno conjunto de genes conservados.
Co-evolução

Interações entre organismos podem produzir tanto conflito e cooperação. Quando a interacção é entre pares de espécies, tais como um agente patogénico e um hospedeiro, ou um predador e sua presa, estas espécies podem desenvolver conjuntos de adaptações combinado. Aqui, a evolução de uma espécie em causa adaptações uma segunda espécie. Estas mudanças em seguida, a segunda espécie, por sua vez, causar novas adaptações nos primeiros espécies. Este ciclo de selecção e de resposta é chamado co-evolução. Um exemplo é a produção de tetrodotoxina no Taricha granulosa e a evolução da resistência tetrodotoxina em seu predador, a serpente de liga comum. Neste par predador-presa, uma corrida armamentista evolutiva produziu altos níveis de toxina na salamandra e correspondentemente elevados níveis de resistência toxina na cobra.
Co-operation
Nem todas as interações co-evoluíram entre espécies envolvem conflito. Muitos casos de interações mutuamente benéficas evoluíram. Por exemplo, existe uma cooperação extrema entre as plantas e os fungos micorrízicos que crescem sobre as raízes da planta e ajudar na absorção de nutrientes do solo. Isto é um relação recíproca como as plantas fornecem os fungos com açúcares da fotossíntese. Aqui, os fungos crescem efectivamente no interior de células de plantas, permitindo a troca de nutrientes com os seus hospedeiros, enquanto que o envio de sinais da planta que suprimem o sistema imunológico .
Coalizões entre organismos da mesma espécie também evoluíram. Um caso extremo é o eusociality encontrado em insetos sociais, como abelhas , cupins e formigas , onde insetos estéreis alimentam e protegem o pequeno número de organismos em uma colônia que são capazes de se reproduzir. Numa escala ainda menor, as células somáticas que formam o corpo de um animal limitar a sua reprodução, de forma que possam manter um organismo estável, que suporta, em seguida, um pequeno número de do animal células germinativas para produzir prole. Aqui, as células somáticas respondem a sinais específicos que eles se instruem a crescer, permanecem como elas são, ou morrem. Se as células ignorar esses sinais e multiplicar de forma inadequada, seu crescimento descontrolado provoca o cancro.
Essa cooperação dentro das espécies podem ter evoluído através do processo de seleção de parentesco, que é onde um organismo age para ajudar a aumentar a prole de um parente. Esta actividade é seleccionado para se porque a ajudar indivíduo contém alelos que promovem a actividade ajudando, é provável que o seu parente irá também conter estes alelos e, assim, esses alelos vai ser transmitida. Outros processos que podem promover a cooperação incluem seleção de grupo, onde a cooperação fornece benefícios para um grupo de organismos.
Especiação

Especiação é o processo em que uma espécie diverge-se em duas ou mais espécies descendentes.
Existem várias maneiras de definir o conceito de "espécie". A escolha de uma definição é dependente das particularidades da espécie em causa. Por exemplo, alguns conceitos de espécie aplicar mais facilmente em direção a organismos de reprodução sexuada, enquanto outros se prestam melhor para organismos assexuados. Apesar da diversidade de vários conceitos de espécie, estes diferentes conceitos pode ser colocado em uma das três abordagens filosóficas amplas: cruzamento, ecológica e filogenética. O conceito biológico de espécie (BSC) é um exemplo clássico da abordagem cruzamentos. Definido por Ernst Mayr em 1942, o BSC afirma que "espécies são grupos de efetiva ou potencialmente acasalando populações naturais, que são reprodutivamente isolados de outros grupos". Apesar da sua ampla utilização e a longo prazo, o BSC como outros não é sem controvérsia, por exemplo, porque estes conceitos não pode ser aplicado aos procariotas, e este é chamado o problema da espécie. Alguns pesquisadores têm tentado uma definição monista unificador da espécie, enquanto outros adotam uma abordagem pluralista e sugerem que pode haver diferentes formas de interpretar logicamente a definição de uma espécie. "
Barreiras à reprodução entre duas populações divergentes sexuais são necessárias para as populações a se tornar novas espécies. O fluxo gênico pode retardar este processo, espalhando as novas variantes genéticas também para as outras populações. Dependendo de quão longe duas espécies divergiram desde o seu ancestral comum mais recente, ele ainda pode ser possível para eles para produzir a prole, como com cavalos e burros que acoplam-se para produzir mulas. Tal híbridos são geralmente inférteis. Neste caso, espécies estreitamente relacionadas podem cruzar regularmente, mas os híbridos serão seleccionados contra e as espécies permanecem distintos. No entanto, híbridos viáveis são ocasionalmente formadas e essas novas espécies podem ter propriedades intermediário entre suas espécies parentais ou possuir um totalmente novo fenótipo. A importância da hibridação na produção de novas espécies de animais não é clara, embora os casos têm sido observados em muitos tipos de animais, com o sapo cinza ser um exemplo particularmente bem estudado.
Especiação foi observado várias vezes sob condições controladas de laboratório e na natureza. Em organismos que se reproduzem sexualmente, resultados de especiação de isolamento reprodutivo seguido de divergência genealógica. Existem quatro mecanismos de especiação. O mais comum em animais é especiação alopátrica, que ocorre em populações geograficamente isoladas inicialmente, como por fragmentação de habitat ou migração. Seleção sob essas condições pode produzir mudanças muito rápidas na aparência e comportamento dos organismos. Como a seleção e deriva ato de forma independente em populações isoladas do resto de sua espécie, a separação pode, eventualmente, produzir organismos que não podem cruzar.
O segundo mecanismo de especiação é a especiação peripatric, que ocorre quando pequenas populações de organismos tornam-se isolado em um novo ambiente. Isto difere da especiação alopátrico em que as populações isoladas são numericamente muito menor do que a população parental. Aqui, o efeito fundador causa especiação rápida após um aumento da consanguinidade aumenta a seleção em homozigotos, levando a mudança genética rápida.
O terceiro mecanismo de especiação é especiação parapátrica. Isto é semelhante ao Peripatric especiação em que uma pequena população entra um novo habitat, mas difere em que não há separação física entre estas duas populações. Em vez disso, especiação resulta da evolução dos mecanismos que reduzem o fluxo de genes entre as duas populações. Geralmente isso ocorre quando houve uma mudança drástica no ambiente dentro do habitat das espécies parentais. Um exemplo é a grama Anthoxanthum odoratum , que podem sofrer especiação parapátrica em resposta a poluição por metais localizada de minas. Aqui, as plantas que possuem resistência a evoluir para níveis elevados de metais no solo. Seleção contra cruzamentos com a população parental sensível ao metal produzido uma mudança gradual no tempo de floração das plantas resistentes de metal, que eventualmente produzidos completo isolamento reprodutivo. Selecção contra híbridos entre as duas populações podem causar reforço , que é a evolução das características que promovem acasalamento dentro de uma espécie, bem como deslocamento de caracteres, que é quando duas espécies tornar-se mais distinta na aparência.

Finalmente, em espécies especiação simpátrica divergem sem isolamento geográfico ou mudanças no habitat. Este formulário é raro, já que uma pequena quantidade de fluxo gênico pode remover as diferenças genéticas entre partes de uma população. Geralmente, especiação simpátrica em animais exige a evolução de ambas as diferenças genéticas e acasalamento não-aleatória, para permitir o isolamento reprodutivo para evoluir.
Um tipo de especiação simpátrica envolve o cruzamento entre duas espécies relacionadas para produzir uma nova espécie híbrida. Este não é comum em animais como animais híbridos são normalmente estéreis. Isso ocorre porque durante a meiose os cromossomos homólogos de cada um dos pais são de espécies diferentes e não pode emparelhar com sucesso. No entanto, é mais comum em plantas de plantas porque muitas vezes duplicar o seu número de cromossomas, para formar poliplóides. Isso permite que os cromossomos de cada uma das espécies parentais para formar pares durante a meiose, uma vez que os cromossomos de cada pai são representados por um par já. Um exemplo de um tal evento de especiação é quando a espécie da planta Arabidopsis thaliana e Arabidopsis arenosa cross-criados para dar a nova espécie Arabidopsis suecica . Isto aconteceu cerca de 20.000 anos atrás, e o processo foi repetido especiação no laboratório, que permite o estudo dos mecanismos genéticos envolvidos neste processo. Na verdade, o cromossomo duplicar dentro de uma espécie pode ser uma causa comum de isolamento reprodutivo, como a metade dos cromossomos duplicados será inigualável ao produzir com organismos undoubled.
Eventos de especiação são importantes na teoria do equilíbrio pontuado , que representa o padrão no registro fóssil de curtas "explosões" de evolução intercaladas com períodos relativamente longos de estase, onde as espécies permanecem relativamente inalterados. Nesta teoria, a especiação ea rápida evolução estão ligadas, com a seleção natural ea deriva genética atuando mais fortemente em organismos submetidos a especiação em novos habitats ou pequenas populações. Como resultado, os períodos de estase no registro fóssil corresponde à população parental e os organismos submetidos a especiação e rápida evolução são encontrados em pequenas populações geograficamente restritas ou habitats e, portanto, raramente sendo preservados como fósseis.
Extinção

Extinção é o desaparecimento de uma espécie inteira. A extinção não é um evento incomum, como espécies aparecem regularmente através de especiação e desaparecem por extinção. Quase todas as espécies de animais e plantas que viveram na Terra estão extintas, ea extinção parece ser o destino final de todas as espécies. Estas extinções ter acontecido de forma contínua ao longo da história de vida, embora a taxa de picos de extinção em massa ocasionais eventos de extinção. O evento de extinção Cretáceo-Paleogene, durante o qual os dinossauros não-aviários foram extintos, é o mais bem conhecido, mas quanto mais cedo Permian evento de extinção -Triassic foi ainda mais grave, com aproximadamente 96% das espécies levadas à extinção. O evento de extinção Holocene é uma extinção em massa contínua associada à expansão da humanidade em todo o mundo ao longo dos últimos milhares de anos. As taxas de extinção atuais são 100-1000 vezes maior do que a taxa de fundo e até 30% das espécies atuais pode ser extinto em meados do século 21. As atividades humanas são agora a principal causa do evento de extinção em curso; o aquecimento global pode acelerar ainda mais no futuro.
O papel de extinção na evolução não é muito bem compreendido e pode depender de qual o tipo de extinção é considerado. As causas dos "de baixo nível" eventos de extinção contínuas, que formam a maioria das extinções, pode ser o resultado da competição entre as espécies de recursos limitados ( exclusão competitiva). Se uma espécie podem competir outra, isso poderia produzir seleção de espécies, com as espécies mais aptos sobrevivem e as outras espécies que estão sendo levados à extinção. As extinções em massa intermitentes também são importantes, mas em vez de agir como força seletiva, eles reduzem drasticamente a diversidade de uma maneira inespecífica e promover rajadas de rápida evolução e especiação em sobreviventes.
História evolutiva da vida
Origem da vida
Química altamente energético é pensado para ter produzido uma molécula auto-replicante ao redor de 4.000 milhões anos atrás, e meio bilhão de anos mais tarde, o último ancestral comum de toda a vida existiu. O actual consenso científico é que o complexo de bioquímica que compõe a vida veio de reações químicas mais simples. O início da vida pode ter incluído moléculas auto-replicantes, tais como RNA e a montagem de células simples.
Descendência comum

Todos os organismos na Terra descendem de um ancestral comum ou conjunto de genes ancestral. Espécies atuais são uma etapa no processo de evolução, com a sua diversidade o produto de uma longa série de eventos de especiação e extinção. O descendência comum dos organismos foi deduzida primeiro de quatro fatos simples sobre organismos: Em primeiro lugar, eles têm distribuições geográficas que não podem ser explicadas pela adaptação local. Em segundo lugar, a diversidade da vida não é um conjunto de organismos completamente única, mas organismos que compartilham semelhanças morfológicas. Em terceiro lugar, características vestigiais com nenhuma finalidade clara assemelham características ancestrais funcionais e, finalmente, que os organismos podem ser classificados usando essas semelhanças em uma hierarquia de grupos aninhados - semelhante a uma árvore genealógica. No entanto, a pesquisa moderna tem sugerido que, devido à transferência horizontal de genes, este " árvore da vida "pode ser mais complicado do que uma árvore de ramificação simples, já que alguns genes se espalharam de forma independente entre espécies de parentesco distante.
Espécies passadas também deixaram registos da sua história evolutiva. Os fósseis , juntamente com a anatomia comparativa dos organismos atuais, constituem o morfológica, anatômica ou, ficha. Ao comparar as anatomias de ambas as espécies modernas e extintas, paleontólogos podem inferir as linhagens dessas espécies. No entanto, esta abordagem é mais bem sucedido para os organismos que tiveram partes do corpo duras, como conchas, ossos ou dentes. Além disso, como procariotas tais como bactérias e archaea partilham um conjunto limitado de morfologias comuns, os seus fósseis não fornecem informações sobre sua ascendência.
Mais recentemente, a evidência para a descendência comum veio do estudo de bioquímicos semelhanças entre organismos. Por exemplo, todas as células vivas usar o mesmo conjunto básico de nucleotídeos e aminoácidos . O desenvolvimento de genética molecular revelou o registro da evolução dos organismos à esquerda em genomas: namoro quando as espécies divergiram através do relógio molecular produzido por mutações. Por exemplo, essas comparações de sequências de DNA revelaram que os seres humanos e os chimpanzés compartilham 98% dos seus genomas e analisar as poucas áreas onde eles diferem ajuda a lançar luz sobre quando o ancestral comum das espécies existiu.
Evolução da vida

Prokaryotes habitado a Terra de aproximadamente 3-4 bilhões de anos atrás. Não ocorreram alterações óbvias em morfologia ou organização celular ocorreu nestes organismos ao longo dos próximos bilhões de anos. As células eucarióticas surgiram entre 1,6-2700000000 anos atrás. A próxima grande mudança na estrutura celular surgiu quando bactérias foram engolidas por células eucarióticas, numa associação cooperativa chamada de endossimbiose. As bactérias encapsulada ea célula hospedeira, em seguida, foram submetidos a co-evolução, com as bactérias a evoluir em mitocôndrias ou hidrogenossomas. Outra imersão de cianobactérias-organismos como, levou à formação de cloroplastos em algas e plantas.
A história da vida foi a dos eucariotas unicelulares procariontes e archaea, até cerca de 610 milhões de anos atrás, quando os organismos multicelulares começaram a aparecer nos oceanos na Ediacaran período. O evolução dos organismos multicelulares ocorreu múltiplas eventos independentes, em organismos tão diversos como esponjas, algas marrons, cianobactérias, micetozoários emixobactérias.
Logo após o aparecimento dos primeiros organismos multicelulares, uma notável quantidade de diversidade biológica apareceu sobre aproximadamente 10 milhões de anos, a um evento chamado explosão cambriana . Aqui, a maioria dos tipos de animais modernos apareceu no registro fóssil, bem como linhagens únicas que posteriormente se tornaram extintas. Vários gatilhos para a explosão cambriana têm sido propostas, incluindo a acumulação de oxigénio na atmosfera a partir da fotossíntese .
Cerca de 500 milhões de anos, plantas e fungos colonizaram a terra e foram logo seguidos por artrópodes e outros animais. Insetos foram particularmente bem sucedidos e até hoje compõem a maioria das espécies animais. anfíbios apareceu pela primeira vez cerca de 364 milhões de anos atrás, seguido pelos primeiros amniotas e aves cerca de 155 milhões de anos atrás (ambos de " réptil ", como linhagens), mamíferos de 129 milhões de anos atrás, Homininae cerca de 10 milhões de anos atrás e os humanos modernos cerca de 250.000 anos atrás. No entanto, apesar da evolução destes animais de grande porte, organismos menores semelhantes aos tipos que evoluíram cedo no processo, continuam a ser altamente bem sucedidos e dominar a terra, com a maior parte de ambas as espécies de biomassa e sendo procariotas.
Aplicações
Conceitos e modelos utilizados na biologia evolutiva, como a seleção natural, têm muitas aplicações.
A selecção artificial é a selecção intencional das características numa população de organismos. Isso tem sido usado por milhares de anos na domesticação de plantas e animais. Mais recentemente, tal selecção tornou-se uma parte vital de engenharia genética, com marcadores seleccionáveis, tais como os genes de resistência a antibióticos ser usadas para manipular o DNA. Em ciclos repetidos de proteínas de mutação e de selecção com propriedades valiosas evoluíram, por exemplo modificado e enzimas novos anticorpos , num processo chamado de evolução dirigida.
Compreender as mudanças que ocorreram durante a evolução do organismo pode revelar os genes necessários para a construção de partes do corpo, os genes que podem estar envolvidos em humanos doenças genéticas. Por exemplo, a tetra mexicano é um albino cavefish que perdeu sua visão durante a evolução. Reprodutores juntos diferentes populações deste peixe cego produziu alguns filhotes com olhos funcionais, uma vez que diferentes mutações havia ocorrido nas populações isoladas que tinham evoluído em diferentes cavernas. Isso ajudou a identificar genes necessários para a visão e pigmentação.
Em ciência da computação , simulações de evolução usando algoritmos evolutivos e vida artificial começou na década de 1960 e foi prorrogado com simulação de seleção artificial. evolução artificial tornou-se um método de otimização amplamente reconhecido como um resultado do trabalho de Ingo Rechenberg na década de 1960. Ele usou estratégias de evolução para resolver problemas complexos de engenharia. Algoritmos genéticos, em particular, tornou-se popular através da escrita de John Holland. As aplicações práticas incluem também a evolução automática de programas de computador. Algoritmos evolutivos são agora usados para resolver problemas multi-dimensionais de forma mais eficiente do que o software produzidos por designers humanos e também para otimizar o projeto de sistemas.
Respostas sociais e culturais

No século 19, especialmente depois da publicação de A Origem das Espécies , em 1859, a idéia de que a vida tinha evoluído era uma fonte ativa de debate acadêmico centrado nas implicações filosóficas, sociais e religiosas de evolução. Hoje, a síntese evolutiva moderna é aceita por uma vasta maioria dos cientistas. No entanto, a evolução continua a ser um conceito controverso para alguns teístas.
Enquanto várias religiões e denominações terem reconciliado as suas crenças com a evolução através de conceitos tais como a evolução teísta, há criacionistas que acreditam que a evolução está em contradição com os mitos de criação encontrados em suas religiões e que levantam várias objeções à evolução. Como já havia sido demonstrado por respostas para a publicação de Vestígios da história natural da criação , em 1844, o aspecto mais controverso da biologia evolutiva é a implicação da evolução humana que os seres humanos compartilham um ancestral comum com os macacos e que as faculdades mentais e morais da humanidade tem os mesmos tipos de causas naturais como outras características hereditárias em animais. Em alguns países, nomeadamente os Estados Unidos, essas tensões entre a ciência ea religião têm alimentado a atual controvérsia criação-evolução, um conflito religioso com foco em política e educação pública. Enquanto outros campos científicos, tais como cosmologia e ciências da Terra também conflito com interpretações literais de muitos textos religiosos , experiências de biologia evolutiva significativamente mais oposição de literalistas religiosas.
O ensino da evolução nas aulas de biologia do ensino secundário americanos era incomum na maior parte da primeira metade do século 20. O decisão Scopes Trial of 1925 fez com que o objeto se torne muito raro em livros de biologia secundárias americanas para uma geração, mas foi gradualmente re- introduzido cerca de uma geração mais tarde e legalmente protegidas com 1968 Epperson v. Arkansas decisão. Desde então, a crença religiosa concorrentes do criacionismo foi legalmente anulado nos currículos do ensino secundário em várias decisões nas décadas de 1970 e 1980, mas retornou em forma pseudocientífica como o design inteligente, a ser excluído uma vez mais em 2005 Kitzmiller v. Dover Area School District caso.